










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new study by Taiwanese researchers from the Academia Sinica and National Taiwan University along with support by scientists from the Medical University of Lublin-Poland and Johns Hopkins University-USA has found that the different SARS-CoV-2 variants not only have different mutations and deletions on them but most importantly they exhibit different but unique spike structures.
The study team reported 38 cryo-EM structures, corresponding to the spike protein of the Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2) and Kappa (B.1.617.1) variants in different functional states with and without its receptor, ACE2.
Interestingly it was found that mutations on the N-terminal domain not only alter the conformation of the highly antigenic supersite of the Delta variant, but also remodel the glycan shield by deleting or adding N-glycans of the Delta and Gamma variants, respectively.
Substantially enhanced ACE2 binding was observed for all variants, whose mutations on the receptor binding domain modulate the electrostatics of the binding interfaces. Despite their abilities to escape host immunity, all variants can be potently neutralized by three unique antibodies.
The study findings were published on a preprint server and are currently being peer reviewed. https://www.biorxiv.org/content/10.1101/2021.09.12.459978v1
The SARS-CoV-2 Variants have been an ongoing concern since their first emergence initially with the Alpha variant or B1.1.7 that was discovered in the UK and which quickly became dominant globally.
Since then, more variants have been discovered, including Beta (B1.351), Gamma (P.1), and the current biggest threat, the Delta variant (B.1.617.2). The Delta variant emerged in India in early 2021 and now accounts for more than 90% of new cases.
The various emerging variants are recognized by mutations of the highly glycosylated spike protein.
This spike protein is largely responsible for the pathogenicity of the SARS-CoV-2 coronavirus. The receptor-binding domain (RBD) within the N-terminal S1 subunit of the spike protein binds to the angiotensin-converting enzyme 2 (ACE2) receptors presented by human cells, allowing viral entry.
Importantly the alterations caused by the mutations to this protein can reduce the effectiveness of treatments such as monoclonal antibodies and lower the effectiveness of vaccines.
It has been found for instance that both the Pfizer and Moderna vaccines have shown up to a 12-fold drop in vaccine effectiveness against the Beta variant.
In order to investigate the impact of these mutations, the study team performed cryo-electron microscopy analysis on the variants spike proteins to determine any structural changes.
The study team examined the structural effect of the mutations in the Beta, Gamma, Delta, and Kappa variants and determined the structures of the 25 spike protein variants.
Importantly it was found that in the Beta variant, the RBD showed three states that were mostly fully open (all RBDs in the up conformation) and two partial ly open, with one or two RBDs in the up conformation. The structure was also solved without any furin cleavage site substitutions or stabilizing tandem profile to confirm the unusual triple RBD-up conformation.
However the Gamma spike protein did not show the same fully open state, only the two partially open states (1 RBD-up and 2 RBD-up), but there were two substates for the 1-RBD-up state when the structure was examined three-dimensionally.
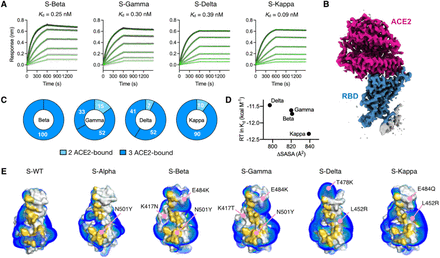
Kinetics and structural basis of receptor ACE2 binding by the SARS-CoV-2 variants. (A) BLI sensorgrams of immobilized ACE2 binding to the S variants. The sensorgrams of the 100, 50, 25, 12.5, 6.25 and 3.125 nM S variants are colored from black to light grey in descending orders superimposed with the fitting results in green. The corresponding Kd values are shown above. (B) Representative cryo-EM map of the RBD S-Delta in complex with ACE2 is colored in blue and magenta for RBD and ACE2, respectively. (C) Distributions of distinct receptor ACE2 binding stoichiometry and quaternary conformations classified according to the number of RBD-bound ACE2. The coloring scheme is shown below. (D) Correlation between the free energy of ACE2 binding and ΔSASA. (E) Surface representations of the RBD of the individual variants with the binding interfaces of the S variants highlighted in gold. Positive isosurfaces of the RBD of the S variants were generated by the APBS module within PyMol. The positions of individual mutations are indicated by pink lines with their changes indicated. The structure of S-WT RBD in complex with ACE2 is taken from the PDB ID 6M0J. The structure of S-Alpha RBD in complex with ACE2 is taken from the PDB ID 7EDJ
Significantly however, the Delta variant showed the most structural variation, including an unusually fully closed conformation in which all RBD’s were in the down conformation. There were also five 1-RBD up substates and three 2-RBD up substates.
Also the most unusual or strange results were seen in the Kappa spike protein, with a previously unseen head-to-head dimer, with all three RBD’s in all three spike proteins binding to another RBD from another spike protein. This structure could be resolved into three substates. Two 1 RBD-up substates were seen, and a single 2 RBD-up conformation and the uncommon all RBD down conformation.
According to the study team, it is thought that these altered conformations allow at least the Gamma and Delta variants to escape immune responses from previous strains, as the chromosomal rearrangements alter the glycan shield around the supersite normally targeted by neutralizing antibodies.
It was also found that almost all of these structural variants also showed greatly enhanced ACE2 binding, with the Kappa variant binding the strongest.
Upon analysis using cryo-electron microscopy, the study team found that the Kappa spike protein dimers showed a reduced negative charge, allowing a much stronger electrostatic binding to the negatively charged ACE2.
Several other mutations in alternate variants had the same effect, to a lesser extent.
The study team had also identified three effective monoclonal neutralizing antibodies against wild-type SARS-CoV-2, the Alpha variant, and D614G, a mutation common to both the Delta and Kappa strains. These are RBD-chAb -15, -25 and -45.
However it was found that RBD-chAb-15 is not effective against the Alpha, Beta, or Gamma variants protected by the N501Y mutation. Still, a mix of the other two antibodies can neutralize both the Alpha variant and variants carrying D614G.
The study findings help further the understanding of the COVID-19 pandemic.
The stud team highlights the benefit of knowing the different structural conformations of the variants and the mechanisms by which they escape immunity from common vaccines and monoclonal antibody treatments.
Importantly the ability of all three discovered neutralizing antibodies to combat the widely spread Delta variant is of great importance, as is the versatility of RBD-chAB-45 in recognizing different states of conformation of the RBD, as it binds to the site la/class 2 on the RBD of the spike protein, which is accessible in either up or down conformations.
Important takeaways from this study include the insight into N-terminal mutations in the variants, which in many cases completely avoid the neutralizing ability of N-terminal domain-specific neutralizing antibodies, further highlighting the importance of targeting the RBD.
Therefore, nAbs that target the RBD remain a preferred route for success, and several RBD-specific nAbs are currently approved for emergency use to treat COVID-19 patients. https://pubmed.ncbi.nlm.nih.gov/34416148/
SARS-CoV-2 genome and mutations
SARS-CoV-2 is a positive-sense single-stranded RNA virus whose genome is of a low stability thus is more prone for mutation accumulation, with approximately 9.8 × 10−4 substitutions/site yearly [3,4,5,6,7]. The architecture of SARS-CoV-2 is made of two groups of proteins: structural proteins (SP) and non-structural proteins (NSP).
SPs are encoded by 4 genes, including E (envelop), M (membrane), S (spike) and N (nucleocapsid) genes [8]. NSPs are mostly enzymes or functional proteins that play a role in viral replication and methylation and may induce host responses to infection.
These genes are encoded in several groups, namely ORF1a (NSP1-11), ORF1b (NSP12-16), ORF3a, ORF6, ORF7a, ORF7b, ORF8 and ORF10.
A variant can be as simply as a virus bearing a deviant mutation or complicated combinations of deviations leading to significant phenotypical alteration from original genome. Although by the beginning of May 2021, there has been reported more than 1.4 million sequences and among them 3913 major representative variants genomes that have been identified and included in the global SARS-CoV-2 sequence database operated by Global Initiative on Sharing Avian Influenza Data (GISAID) [9], not all genetic mutations lead to variation in major proteins and/or alter virus infectivity. The spike gene mutations account for most of the clinically influential VOCs while the ORF1a frame of the genome serves as a key region for NSP mutations.
We will focus our discussion here on the VOCs that have major global health impacts since the 4th quarter of the year 2020, including Alpha variant (B.1.1.7), Beta (B.1.351), Gamma (P.1) and Kappa and Delta (B.1.617.1 and B.1.617.2) (Table 1).
Spike mutations
Spike protein mediates the virus attachment to human cell surface angiotensin converting enzyme 2 (ACE2) receptor, thus facilitating viral entry during infection [10,11,12]. It is split into two subunits, S1 and S2. The S1 unit possess the receptor-binding domain (RBD) which can directly bind to ACE2 receptor and is also the dominant target of neutralizing antibodies (Ab) against SARS-CoV-2. S1 is thus considered a hotspot for mutations that may have high clinical relevance in terms of virulence, transmissibility, and host immune evasion [13,14,15,16] (Table 2).
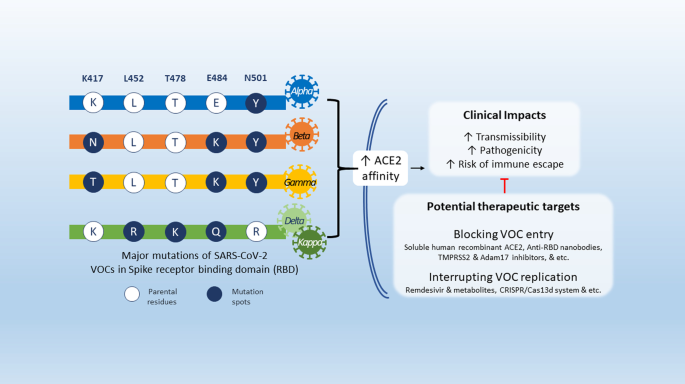
The Alpha variant has an N501Y mutation: at the 501 residue, N asparagine has been replaced with Y tyrosine, as well as K417N—lysine K replaced with asparagine N [9]. An emerging variant derived from B.1.1.7 also carries E484K mutation—glutamic acid E replaced with lysine K [9]. Both Beta and Gamma variants have more substitutions other than N501Y [9]. The Beta variant has E484K, while the Gamma variant has the E484K and the K417T mutations [9].
The latest major variants, Delta and Kappa, sharing two mutations E484Q (glutamic acid E substituted by glutamine Q) and L452R (leucine L altered by arginine R) were identified in India’s second COVID-19 wave. Other than the two mutations above, Delta also harbours a unique mutation, T478K (threonine T replaced by lysine K) [9].
The S1 mutations significantly increases the binding affinity to ACE2 while showing lower affinity to neutralizing antibodies [17,18,19,20,21], suggesting a possible explanation for their occurring higher transmissibility and virulence [22, 23].
Another mutation at non-RBD sites, named D614G, is the most spreading mutation carried by over 99% of prevalent variants since early 2020 [23, 24]. Such mutation does not change the binding affinity to ACE2 or neutralizing Abs for the virion, yet it may increase spike density by preserving the integrity of spike and avoiding S1 shedding [25]. With more functional spikes available, D614G variants are armed with increased infectivity and hence increased replication in vitro while earlier transmission in vivo [23, 25, 26].
Recently, increasing deletions are observed in the neutralizing Ab-recognizing domain, namely recurrent deletion regions (RDRs), in the N-terminus of S1 subunit [27]. Deletions in RDRs wipe out the epitopes, and eventually aiding the virus evading host’s immune supervision and potentially defecting certain neutralizing Abs or vaccines. A majority of Alpha derived variants (ΔRDR1, S: ΔHV 69–70, & ΔRDR2, S: ΔY144), Beta derived variants (ΔRDR4, S: ΔLAL 242–244) and B.1.36 (ΔRDR3, S: ΔI210) carry this kind of mutation [27].
NSP mutations
Two mutation hot-spots, NSP1 of ORF1a/ORF1ab, and ORF8, have been found related to the virulence and transmissibility. NSP1 is a key protein to antagonize type I interferon induction in the host and benefit the replication of the virus itself [28, 29]. ORF8 is known as an immune-evasive protein that downregulates major histocompatibility complex class I (MHC-I) in host cells [30, 31]. Recently, the Alpha variant, identified from a single immunocompromised individual, was shown to contains a premature stop codon at position 27 of ORF8[32].
Variants with partial deletion of NSP1 and ORF8 have been identified (e.g., the NSP1: Δ500-532 variant in Sichuan, China, and the ORF8: Δ382 variant in Singapore) [29, 31]. Despite that truncated NSP1 and ORF8 both contribute to milder infections [29,30,31] and account for less than 5% of infections worldwide, they have become the major variants in Africa since late 2020[9].
Potential clinical impacts of SARS-CoV-2 VOCs
Increased transmissibility and viral virulence
It was shown that S-protein mutation D614G may impact SARS-CoV-2 transmissibility rate due to higher affinity for olfactory epithelium and it was shown to have higher transmissibility in animal models [33, 34]. It was also shown that it has a higher virion stability and was shown to be more resistant to proteolytic cleave as well as higher viral titer in upper airways [35, 36] suggesting that it may potentially affect virus transmissibility and virulence. Yet, it showed increased susceptibility for neutralizing antibodies and no difference in clinical severity nor hospitalization outcomes and mortality was observed [37, 38].
Evidence suggest that the VOCs Alpha and Beta increased transmissibility rate at ~ 50% especially in younger group ages and children [39, 40]. Alpha variant was shown to increase hospitalizations and mortality that may be attributed to their escape from neutralizing Abs due to their RBD mutations [41].
The Epsilon variant (B.1.427/B.1.429, California variants) increased transmissibility up to 24% with higher viral shedding, which is attributed to the of L452R spike mutation that was shown to stabilize spike-ACE2 receptor interaction [42, 43].
Although it is also suggested that other variants such as Gamma, Epsilon variants and recent Iota variants (B.1526, New York variant) may also have increased virulence due to spike mutations that increase affinity to ACE2, there is still no data available regarding viral virulence.
Decreased diagnostic sensitivity
The new VOCs can reduce the detection sensitivity of RT-PCR based diagnostic tools especially when mutations occur in locations where probes and primers may bind [44]. Reports suggest that 79% of the primer binding sites used in the RT-PCR assay are already mutated in at least one genome with the highest significance of the GGG → AAC substitution [45]. Recent analysis which mapped primers or probes binding sites showed a cumulative variants frequency of ≥ 1% in the global SARS-CoV-2 genomes [46].
The Alpha lineage was shown to have higher false-negative results when using specific commercial kits directed to the spike (S) gene but not when using standard protocols such as Berlin-Cherite protocol since it does not involve the S protein-encoding gene as target [47].
Another concern is a variant detected in France of a S deletion (ΔH69-V70) which has shown to be associated with S-gene target gene detection failure in three-target RT-PCR [48]. Several reports have targeted mutations in different open reading frames (ORFs) especially ORF8 position which was found in some isolates from Mexico, Belize and Guatemala as potentially leading to epitope loss and reduced sensitivity for serological testing [49,50,51].
On the other hand, other studies showed that although mismatches in the primer/probes binding regions of SARS-CoV-2 diagnostic assays can be detected in different SARS-CoV-2 variants, they were tolerated and did not result in reduced assay performance and false-negative results [52, 53]. Moreover, according to bioinformatic analysis performed, the known variability occurring in the SARS-CoV-2 population have minimal or no effect on the sensitivity existing diagnostic tools for viral detection [54, 55].
Still, the continuous emergence of SARS-CoV-2 variants and possible mismatches highlight the importance of global molecular surveillance and designing diagnostic strategies such as combining diagnostic methods during future outbreaks or perform assays that target two or more positions in highly conserved regions of the viral genome to promote higher specificity and sensitivity results as well as developing highly specific diagnostic tools using CRISPR [56, 57].
Potential influence on vaccination
Currently, all vaccines are based on introducing spike protein, which is the major superficial virulence of SARS-CoV-2, using the reference genome isolates early in the pandemic. As there is no sufficient evidence to support the effect of vaccines against Delta and Kappa variants, we’ll focus on the Alpha, Beta and Kappa variants.
mRNA Vaccines
Two major mRNA-based anti-SARS-CoV-2 vaccines have been approved: BNT162b2 (Pfizer-BioNTech) and mRNA-1273 (Moderna). Studies suggest that BNT162b2 vaccines were able to stimulate the recipients to generate capable antibodies to neutralize Alpha and Gamma variants yet being significantly less protective against Beta variant [41, 58, 59] mRNA-1273 was shown to enhance sufficient neutralizing ability against Alpha variant yet lower reciprocal titer against Beta variant [41, 60, 61].
Adenovirus-based vaccines
There are 4 adenovirus-based vaccines that have been authorized for general or emergency use. Among of which, Ad26.COV2.S is a recombinant, replication-incompetent adenovirus serotype 26 (Ad26) vector encoding a full-length and stabilized SARS-CoV-2 spike protein (Janssen) was shown to have reduced efficacies to Beta variant (64%) and Gamma dominant Latin America variant (61%), compared to the U.S. (72%) where Alpha is the dominant strain [62].
ChAdOx1 nCoV-19 (Oxford) is a chimpanzee adenovirus-vectored vaccine expressing the SARS-CoV-2 spike protein. Recent studies revealed that the efficacy of ChAdOx1 nCoV-19 was 74.6% against Alpha but as low as 10.4% against Beta [63, 64]. Gam-COVID-Vac (Ad26 and Ad5) is also claimed protective to the global VOCs, yet the clinical trial result has not yet been publicized [65].
An ongoing clinical trial on the combination of ChAdOx1 nCoV-19 and Gam-COVID-Vac (Russia), which is a heterologous COVID-19 vaccine consisting of two components, a recombinant adenovirus type 26 (rAd26) vector and a recombinant adenovirus type 5 (rAd5) vector, both carrying the gene for SARS-CoV-2 spike glycoprotein. There is no data regarding its efficacy on VOCs [66].
Subunit vaccines
NVX-CoV2373 (Novavax) contains a full-length, prefusion spike protein, and shows an 86.3% efficacy against Alpha, yet 48.6% against Beta [67]. However, none of the recombinant protein-based vaccines have yet to be approved for general use.
Inactivated virus-based vaccines
Three inactivated virus- based vaccines have been approved so far and have been widely used in China, India and Brazil. A recent in vitro study suggests that antisera elicited by BBIBP-CorV vaccine (Sinopharm) are able to neutralize the Beta variant in a differentially weaker level compared to the wildtype strain and the D614G variant [68].
A recent serological study has shown that BBV152 (Bharat Biotech International Limited) vaccinated human serum is able to neutralize the Alpha variant [69]. Preliminary data from a study conducted in Sao Paulo, Brazil indicate that the most widely vaccinated vaccine, CoronaVac (Sinovac Biotech), is effective against Gamma variant [70]. The same research facility claimed the vaccine also ‘works well’ against the Alpha and Gamma variants [71].
To conclude, it appears that Beta is most likely variant to affect the approved vaccines efficiency while Alpha and Gamma variants do not. These results suggest that a new vaccine might be required specifically to target Beta variant. Many strategies are currently under development to cope with Beta variant challenge such as booster vaccines [72].
Potential therapeutics for VOCs
S1 RBD targeted therapy
Whether specifically targeting spike proteins using small peptide-based therapies or using single-domains neutralizing antibodies against any of those targets, these therapeutic strategies efficiency may be compromised by the emergence of SARS-CoV-2 variants especially those possessing spike proteins and RBD mutations that increase affinity to ACE2 such as Alpha, and Iota variant, by potentially escaping neutralizing antibodies and competing with those agents for the same binding targets [73,74,75].
In order to avoid antibody escape, strategies to combine different neutralizing antibody cocktail have been suggested as a therapeutic approach against the emerging variants [76]. Other treatments such as anti-RBD nanobodies isolated from llamas were shown to neutralize RBD variants suggesting they might be a promising tool against new SARS-CoV-2 VOCs as well [77, 78].
Different engineered variants of human recombinant soluble ACE2 (hrACE2), were reported to significantly inhibit SARS-CoV-2 infection in vitro and causing sustained viral entry blockade upon engagement of hrACE2 with the RBD in SARS-CoV-2 S protein with high affinity [79,80,81]. This is a potentially powerful treatment against SARS-CoV-2 VOCs as it can exploit the increase S-protein host receptor-binding affinity caused by S-mutations, toward increasing S-protein affinity to hrACE2. Moreover, no mutations that limit receptor-binding affinity were discovered as this will decrease affinity to native ACE2 receptor and may likely to attenuate virulence [82], suggesting that viral escape from hrACE is very unlikely.
Interruption of endosomal formation
Targeting endosomal formation of SARS-CoV-2 to block entry to host cells such as antimalarial drugs and macrolides, and us of drugs targeting host cell transmembrane protease serine 2 (TMPRSS2) such as Camostat [83,84,85] or A disintegrin and metalloprotease 17 (ADAM17) inhibitors [86].
Interruption of SARS-CoV-2 VOC genome
Promising antiviral drugs such as the FDA-approved Remdesivir and its metabolites, Ribaverin and Galidesivir have been shown to inhibit viral replication in vitro and in vivo studies due to their effect on inhibiting RNA dependent RNA polymerase (RdRp) [87, 88]. The discovery of RdRp hotspot mutations in SARS-CoV-2, found mostly in European strains may lead to drug-resistance of to RdRp inhibitors in a similar mechanism found in Influenza and Hepatitis C [89,90,91]. However, it has been shown currently that those variants have minimal impact for pre-existing resistance to Remdesivir.
Another potential approach is Prophylactic Antiviral CRISPR in Human Cells (PAC-MAN), which is a Cas13d-based strategy that target reserved regions such as nucleocapsid protein and RdRp in SARS-CoV-2 viral genome and may serve as pan-coronavirus strategy for any future coronaviruses and variant that may emerge [92].
reference link : https://ccforum.biomedcentral.com/articles/10.1186/s13054-021-03662-x



{kind=link}