










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



The phytochemical erianin, is a natural plant-derived bibenzyl compound, present in Dendrobium chrysotoxum, an orchid species native to Southeast Asia.
It has been found that in the early stages, the development of prostate cancer is dependent on androgens. Over time with androgen deprivation therapy, 20% of prostate cancers progress to a castration-resistant form.
The study team reports herein that erianin exhibits anti-tumor effects in androgen-sensitive and castration-resistant prostate cancer cells through different mechanisms.
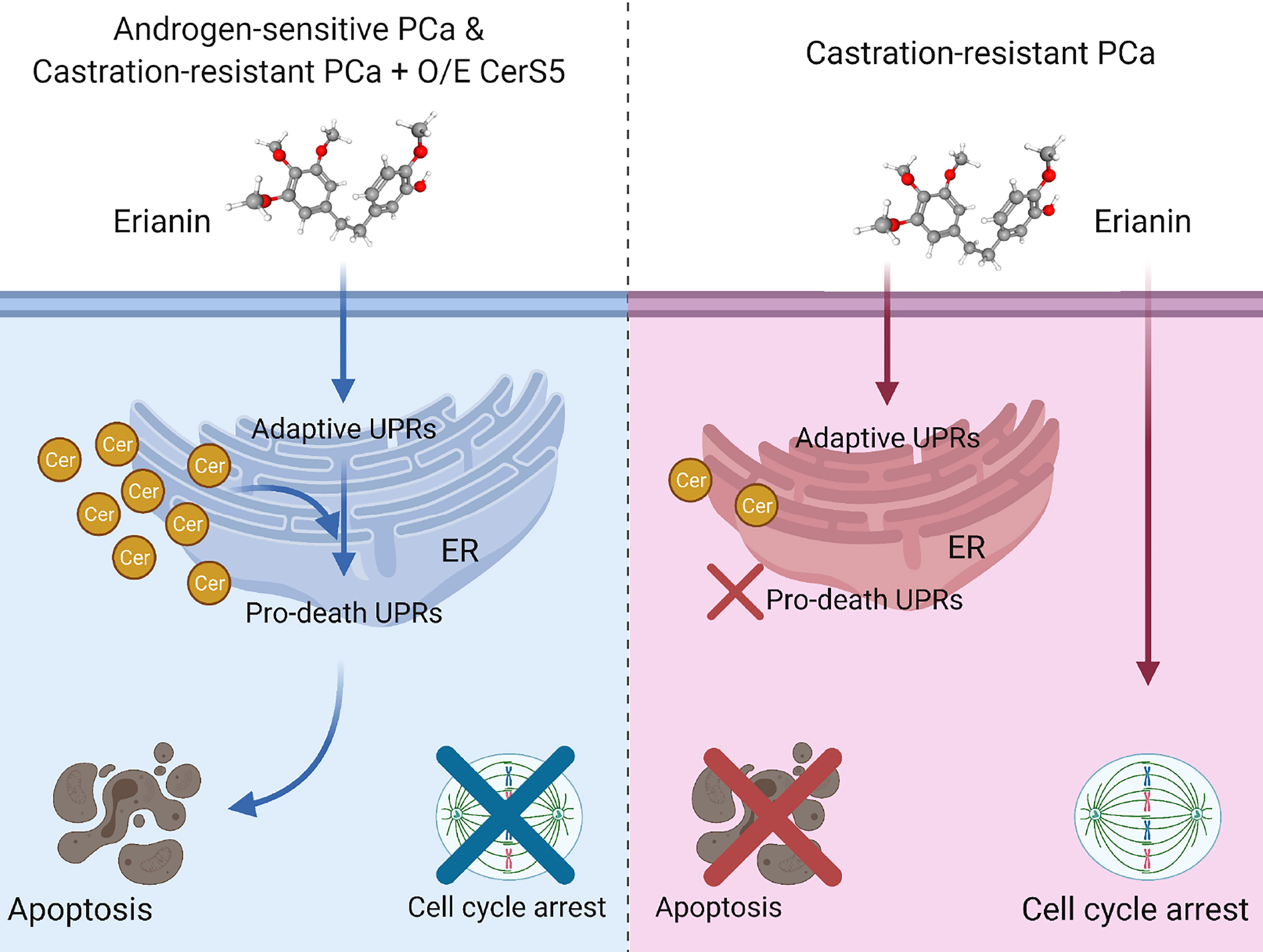
The study found that erianin induces endoplasmic reticulum stress-associated apoptosis in androgen-sensitive prostate cancer cells. It also triggers pro-survival autophagic responses, as inhibition of autophagy predisposes to apoptosis.
However in contrast, erianin fails to induce apoptosis in castration-resistant prostate cancer cells. Instead, it results in cell cycle arrest at the M phase. Mechanistically, C16 ceramide dictates differential responses of androgen-sensitive and castration-resistant prostate cancer cells to erianin.
Erianin elevates C16 ceramide level in androgen-sensitive but not castration-resistant prostate cancer cells. Overexpression of ceramide synthase 5 that specifically produces C16 ceramide enables erianin to induce apoptosis in castration-resistant prostate cancer cells.
The study findings provide both experimental evidence and mechanistic data showing that erianin is a potential treatment option for prostate cancers.
The study findings were published in the peer reviewed journal: Frontiers in Oncology. https://www.frontiersin.org/articles/10.3389/fonc.2021.738078/full
Prostate cancer is the second most common cancer diagnosis and the third leading cause of cancer death in American men.
According to the American Cancer Society, an estimated 191,930 men were newly diagnosed with prostate cancer in 2020. It is also estimated that there will be 33,330 prostate cancer related deaths this year alone.
It was also reported that 1 in 9 American men will receive a diagnosis of prostate cancer in their lifetime!
Although prostate cancer is very common, it is slow growing, meaning most men diagnosed with prostate cancer will not die from it.
According to the latest Cancer News, prostate cancer is also the sixth highest cause of cancer-related mortality worldwide.
The study team found that erianin had anti-tumor effects on both androgen-dependent (early-stage) and castration-resistant (advanced-st age) prostate cancer cells.
Dr Yanfei (Jacob) Qi, Head of the Lipid Cell Biology Laboratory at the Centenary Institute at the University of Sydney and lead researcher on the study told : “Early in their development, prostate cancers need androgens (male sex hormones), including testosterone, to grow.
Androgen deprivation therapy, also known as hormone therapy, aims to decrease androgen levels and can help slow or limit the cancer’s growth.
When the prostate cancer stops responding to this treatment and continues to grow, the prostate cancer is at an advanced stage known as castration-resistant.”
The study findings showed that erianin elevated levels of a fatty acid called C16 ceramide inside the androgen-dependent prostate cancer cells. This caused the cells to die through a process called endoplasmic reticulum stress-associated cell death.
However in contrast, erianin alone failed to elevate C16 ceramide levels in the castration-resistant prostate cancer cells. However, artificially increasing C16 ceramide in these cells, in conjunction with the use of erianin did result in successful cell death.
Dr Qi added, “Novel treatments for prostate cancer are urgently needed. Up to twenty percent of patients receiving androgen deprivation therapy progress to advanced prostate cancer within five years.
We’ve shown that erianin could play an important role in the development of new medical drugs that are able to target both early and late-stage prostate cancers, potentially benefiting many patients and helping save lives.”
Another study has shown that erianin is able inhibits lung cancer cell growth and migration via calcium/calmodulin-dependent ferroptosis.
https://www.nature.com/articles/s41392-020-0149-3
It has also shown to be able to inhibit cervical cancer cells.
https://www.spandidos-publications.com/10.3892/ol.2018.9267
In fact its application in the field of cancer treatments is gradually recognition.
There are numerous studies showing its anticancer properties.
https://www.dovepress.com/progressive-study-of-effects-of-erianin-on-anticancer-activity-peer-reviewed-fulltext-article-OTT
https://www.frontiersin.org/articles/10.3389/fonc.2021.727605/full
https://portlandpress.com/bioscirep/article/41/6/BSR20210093/228754/Erianin-induces-triple-negative-breast-cancer
https://www.worldscientific.com/doi/10.1142/S0192415X2050010X
Prostate cancer (PCa) is the second most common cancer and the sixth highest cause of cancer-related mortality worldwide (1). In the United States, PCa accounts for 26% and 11% of all cancer incidence and death in men, respectively (2). In the early stages of development, PCa relies on androgen to stimulate its growth, and thus it is sensitive to androgen deprivation therapy (3, 4).
However, 20% of patients receiving androgen deprivation therapy progress to castration-resistant PCa (CRPC) within 2-3 years. CRPC is associated with a significantly higher degree of drug resistance (5). Meanwhile, androgen deprivation therapy has shown adverse effects, including cardiovascular, metabolic and cognitive morbidities (4).
Taxanes, such as docetaxel and cabazitaxel, are applied as primary chemotherapy in CRPC patients (3, 4, 6). These chemicals can disrupt microtubule dynamics during both mitosis and interphase, leading to cell cycle arrest and even death in PCa (6, 7). However, approximately half of patients treated with taxanes develop drug resistance (8).
The prognosis and overall survival rate of PCa remain unsatisfactory, and searching for new treatment options is still the key matter in PCa research (6).
Natural products are valuable sources of both new anti-cancer medicines and lead compounds (9). Bibenzyls are a class of phenolic compounds generated as ethane derivatives from the flavonoid biosynthetic pathway in plants (10, 11). Among the 89 bibenzyls identified from the Dendrobium genus, 23 compounds exhibit anti-tumor properties by inducing apoptosis, inhibiting cell proliferation and suppressing cell migration and invasion in multiple cancer cell lines [summarized in (11)].
Erianin is a major bibenzyl compound extracted from Dendrobium. This phytochemical has little toxicity in immortalized normal human hepatocytes and nasopharyngeal normal primary cells (12, 13), whereas it has shown potent anti-tumor effects at nanomolar concentrations.
Erianin induces oxidative stress, leading to apoptosis via the intrinsic apoptosis pathway (14–18), blocks the cell cycle at the G2/M phase (12, 14, 17), and has been recently found to trigger ferroptosis, an iron-dependent programmed cell death (19).
In addition, treatment of nude mice with erianin significantly inhibit the growth of lung cancer, osteosarcoma, colorectal cancer and bladder cancer xenografts (14, 16, 17, 19–21). However, the anti-tumor effects of erianin in PCa have not been examined.
Endoplasmic reticulum (ER) stress-associated apoptosis is one of the primary targets of anti-cancer drug discovery (22), and a large number of bioactive natural products elicit anti-tumor activities in this manner (23, 24). The ER is an intracellular membrane network, maintaining proteostasis within the cell.
Perturbance of ER homeostasis will activate unfolded protein responses (UPRs) through three canonical signaling axes, including PKR-like ER kinase (PERK)-eukaryotic initiation factor 2α (eIF2α), inositol-requiring enzyme 1α (IRE1α), and activating transcription factor 6 (ATF6), leading to adaptation to the stress (22, 25).
In response to severe and prolonged stress, all three signaling arms activate pro-apoptotic UPRs, converging at the induction of C/EBP homologous protein (CHOP) (25). CHOP transcriptionally upregulates Bim, a pro-apoptotic Bcl-2 family member, which is believed to be the mechanistic link between ER stress and the intrinsic apoptosis pathway (26).
Erianin induces phosphorylation of c-Jun N-terminal kinases (JNK), a signaling node associated with both ER stress and oxidative stress (14, 16, 18). However, a definitive answer to whether erianin provokes ER stress-associated apoptosis still remains elusive.
Autophagy is another adaptive response of cells to cope with stress (27). Upon the activation of autophagy, pro-light chain 3 (LC3) is cleaved in the cytosol to form LC3-I, followed by the lipidation of LC3-I to yield LC3-II (28). LC3-II is localized to autophagosomes and drives their maturation (28).
After the fusion of autophagosomes and lysosomes, cargoes contained in autophagosomes, such as p62, are degraded along with LC3-II in lysosomes (29). Highly dependent on the context, autophagy can be either pro-survival or pro-death, as a double-edged sword. In addition, chemotherapies often induce autophagy in parallel with apoptosis (30, 31).
Thus, although autophagy is a popular target in anti-cancer clinical trials, care must be taken to understand the role of autophagy in the context of the specific disease state (32). Erianin has been demonstrated to induce pro-survival autophagy in human osteosarcoma cells (14), making it intriguing to examine the role of erianin-induced autophagy in PCa.
Sphingolipids are a class of essential lipids, functioning as both membrane constituents and signaling molecules within the cell (33). Metabolically, sphingolipids are interconnected in a network with ceramide as the central hub (33). Ceramide can be biosynthesized from the condensation of amino acids and fatty acids, followed by reduction, acylation and desaturation (33). Ceramide can be degraded via the sphingolipid catabolic pathway into non-lipid products (33).
In ceramide biosynthesis, ceramide synthases (CerS) mediate the rate-limiting step that adds the fatty acyl chain to the sphingoid base (34). There are six mammalian isoforms of CerS, designated as CerS1-6 (34). Of them, CerS5 and CerS6 predominantly produce C16 ceramide (35). In the prostate, CerS5 is more prevalent than CerS6 (36).
Ceramide orchestrates apoptosis, ER stress and autophagy signaling (37, 38). However, different ceramide species may exhibit distinct biological roles. For example, C16 ceramide is a well-established pro-apoptotic signal, whereas C24 ceramide is often anti-apoptotic (39–41).
In addition, C16 ceramide has different biological effects at spatially distinct subcellular compartments. For instance, ablation of CerS6 but not CerS5 confers metabolic protection in the liver, as CerS6 mainly produces C16 ceramide in mitochondria (42). So far, research on the role of ceramide in prostate cancer has not reached the level of the subcellular lipid pool or lipid subtype.
Instead, total ceramide levels have been found to determine therapy resistance in PCa cells. The anti-cancer drug camptothecin elevates total ceramide mass and thus induces apoptosis in androgen-sensitive LNCaP PCa cells, but it fails to do so in castration-resistant PC3 PCa cells (43). Accumulation of ceramide by blocking its catabolism potentiates the apoptosis of PC3 cells (44, 45). However, whether erianin regulates ceramide remains untested.
In this study, we assessed the anti-tumor properties of erianin, including its effects on clonogenicity, cell migration and cell viability in LNCaP and PC3 cells. To elucidate the anti-tumor pathways induced by erianin, we examined ER stress-associated apoptosis and cell cycle arrest. Here we identified that erianin elicits discrepant anti-tumor mechanisms in LNCaP and PC3 cells.
We also examined whether erianin regulated ceramide levels in PCa cells and elucidated whether C16 ceramide determines PCa cell susceptibility to erianin-induced apoptosis. Collectively, our study aims to demonstrate the potential therapeutic benefits of erianin against PCa and elucidates the underlying anti-tumor mechanism.
REFERENCE LINK : https://www.frontiersin.org/articles/10.3389/fonc.2021.738078/full



{kind=link}