










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Un nuovo studio condotto da ricercatori francesi dell’Institut Pasteur-Paris, dell’Université de Paris, del Vaccine Research Institute-France e della Sorbonne Université-Paris ha scoperto in modo allarmante che le varie nuove varianti emergenti di SARS-CoV-2 mostrano una maggiore formazione di sincizi.
La sincizia è formata dalla fusione di cellule infette con cellule vicine che portano alla formazione di cellule ingrandite multinucleate.
Questo evento è indotto dall’espressione superficiale della proteina di fusione virale che è fusogena direttamente sulla membrana della cellula ospite. La sincizia può verificarsi solo con virus in grado di fondersi direttamente sulla superficie cellulare senza la necessità di endocitosi.
Tipicamente, il COVID-19 grave è caratterizzato da anomalie polmonari, inclusa la presenza di pneumociti sinciziali.
Il potenziale di formazione di sincizi delle proteine varianti spike rimane scarsamente caratterizzato.
Il team di ricercatori ha prima valutato la diffusione e la fusione di Alfa (B.1.1.7) e Beta (B.1.351) nelle colture cellulari, rispetto al ceppo ancestrale D614G. Alfa e Beta si sono replicati in modo simile al ceppo D614G in Vero, Caco-2, Calu-3 e nelle cellule delle vie aeree primarie.
Tuttavia, Alpha e Beta formavano sincizi più grandi e più numerosi. Le proteine spike varianti hanno mostrato una maggiore affinità ACE2 rispetto a D614G.
La fusione alfa, beta e D614G è stata similmente inibita dalle proteine transmembrana indotte dall’interferone (IFITM). Le singole mutazioni presenti nei picchi Alpha e Beta hanno modificato la fusogenicità, il legame con ACE2 o il riconoscimento da parte degli anticorpi monoclonali.
I risultati dello studio sono stati pubblicati nell’EMBO Journal, sottoposto a revisione paritaria.
collegamento di riferimento: https://www.embopress.org/doi/abs/10.15252/embj.2021108944
I sincizi sono strutture cellulari evolutivamente conservate formate dalle fusioni cellulari multiple di cellule uninucleari. Nei mammiferi, il miglior esempio di sincizio fisiologico sono le fibre muscolari, che contengono migliaia di cellule muscolari fuse per consentire la loro rapida contrazione coordinata [7].
È anche importante nel processo di decidualizzazione durante l’impianto dell’embrione [8]. La sincizia può anche essere indotta da alcuni tipi di infezioni da virus, come il virus dell’immunodeficienza umana, il virus respiratorio sinciziale e il virus dell’herpes simplex [9].
Si potrebbe immaginare che la fusione cellulare indotta da virus faciliti il trasferimento dei genomi virali alle cellule vicine. Tuttavia, i meccanismi virali e cellulari che regolano la formazione di sincizi durante l’infezione da SARS-CoV-2 rimangono in gran parte sfuggenti.
Durante l’esame delle sezioni polmonari istopatologiche di pazienti deceduti per COVID-19, il gruppo Giacca e il gruppo Sun hanno osservato l’esistenza prevalente di cellule atipiche contenenti 2-20 nuclei.
Le identità di questi sincizi sono state successivamente confermate dalla loro espressione di produttori specifici di pneumociti. Il test di co-coltura in vitro ha mostrato che la linea cellulare epiteliale del rene di scimmia, le cellule Vero (ACE2+), dopo aver espresso la proteina spike SARS-CoV-2, potrebbe formare sincizi omologhi o fondersi con altre linee cellulari purché fosse presente il recettore ACE2.
È interessante notare che quando le cellule Vero sono state trasfettate con la proteina Spike di SARS-CoV-1, non è stata osservata alcuna formazione di sincizi. Pertanto, l’elemento chiave responsabile della sincizia mediata da SARS-CoV-2 è assente nella proteina spike di SARS-CoV-1. Spinti da questa ipotesi, Sun et al. ha confrontato la proteina spike di SARS-CoV-2 e SARS-CoV1 e ha scoperto che c’è un inserimento di quattro amminoacidi (PRRA) prima del sito di scissione S1/S2 nella proteina spike SARS-CoV-2.
La mutazione troncata della proteina spike SARS-CoV-2 con delezione “PRRA” perde la sua capacità di fondere le cellule. Coerentemente, la proteina spike di SARS-CoV1 ha indotto efficacemente il sincizio una volta che la sequenza “PRRA” è stata inserita prima del sito di scissione S1/S2 del genoma SARS-CoV-1.
Inoltre, il gruppo Sun ha dimostrato che un motivo bi-argininico contenente R682 e R685 determina la formazione del sincizio costruendo mutazioni singole o combinate nell’inserzione “PPRA”. Fig. 1.
Fig. 1: La proteina spike SARS-CoV-2 e il canale ionico cellulare TMEM16 hanno mediato in modo collaborativo la formazione di sincizi nelle infezioni da COVID-19.
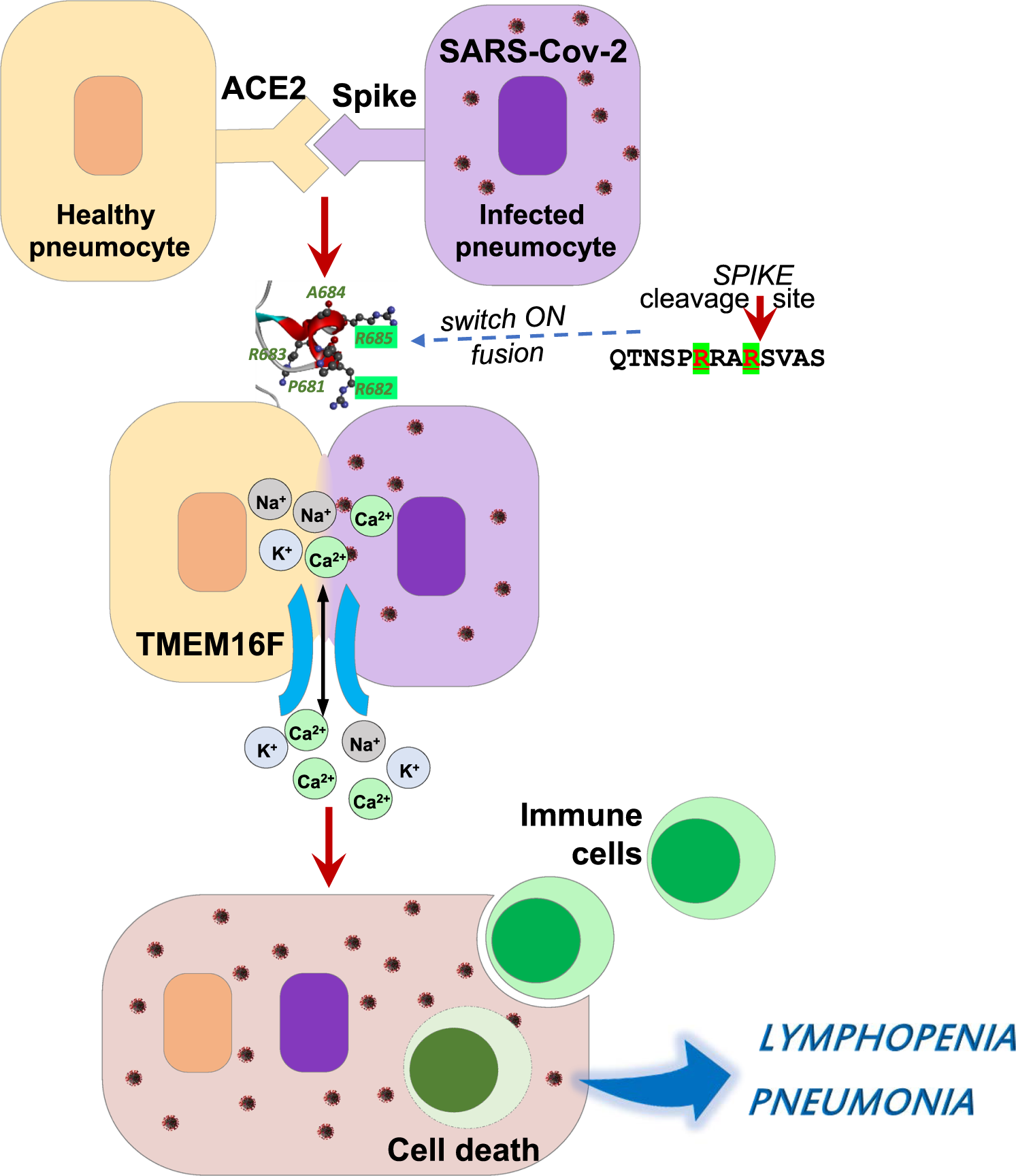
I nuovi dati ottenuti da Sun et al. forniscono informazioni critiche per comprendere la sincizia attraverso la decifrazione della struttura di base richiesta per la fusione cellulare mediata dalla proteina spike di SARS-CoV-2, mentre Giacca et al. focalizzato sul meccanismo cellulare e sul potenziale terapeutico della sincizia durante l’infezione da SARS-Cov-2.
A questo proposito, Giacca e colleghi hanno esaminato 3049 farmaci approvati dalla FDA/EMA utilizzando la proteina spike SARS-CoV-2 che esprime il sistema di fusione cellulare in vitro basato su cellule Vero per cercare farmaci che bloccano la sincizia.
È interessante notare che i farmaci che hanno soppresso la fusione cellulare sono tutti in grado di regolare i livelli intracellulari di Ca2+. Tra i farmaci bloccanti la sincizia, la niclosamide, un agente antielmintico orale, è risultata efficace a una dose molto bassa (IC50 = 0,34 μM) e potrebbe prevenire la morte cellulare indotta da virus.
La niclosamide è un potente antagonista della famiglia dei canali del cloro attivati da Ca2+ TMEM16/anoctamina [10]. TMEM16F è stato anche notevolmente aumentato nelle cellule Vero dopo l’espressione della proteina spike. Quando l’espressione di TMEM16F è stata disturbata, la formazione di sincizi nelle cellule con espressione di picco è stata ridotta. Pertanto, l’attivazione di TMEM16F è il segnale responsabile dell’attivazione della sincizia.
Questi due eleganti studi hanno rivelato collettivamente un nuovo concetto di formazione di sincizi e i suoi ruoli nelle infezioni da SARS-CoV-2, che possono essere brevemente precisati come segue. Le infezioni da SARS-CoV-2 inducono l’espressione superficiale della glicoproteina spike.
L’interazione della proteina spike con il recettore ACE2 delle cellule vicine attiva quindi TMEM16F e innesca l’insalubrità del frammento S2 della profusione della proteina spike in modo dipendente dal motivo bi-arginina, che alla fine porta alla fusione della membrana e alla formazione di sincizi . Tuttavia, ci sono ancora molte domande che restano da chiarire.
Uno dei quali è se il motivo bi-arginina è richiesto per l’attivazione di TMEM16F. Un altro è l’impatto della formazione di sincizi sulle infezioni da SARS-CoV-2 in vivo. Sole et al. hanno scoperto che un tipo di struttura cellulare positiva per CD45 si presenta nei sincizi dei pazienti COVID-19.
Potrebbe trattarsi di una struttura cellula nella cellula. Quando le cellule mononucleate del sangue periferico umano sono state co-coltivate con sincizi indotti dalla proteina spike SARS-CoV-2, potrebbero essere inghiottite e morire all’interno del sincizio, fornendo così una possibile spiegazione per la linfopenia nelle infezioni da SARS-CoV-2 [11] .
Si può altamente sospettare che i sincizi siano deleteri per i pazienti COVID-19 poiché i sincizi sono stati osservati solo nelle fasi gravi delle malattie e i sincizi possono indurre linfopenia. Nonostante l’osservazione di pneumociti multinucleati nell’autopsia, non è ancora noto se tali sincizi svolgano un ruolo critico nella patogenesi delle CRD di pazienti gravi con COVID-19. Recentemente, in uno studio clinico randomizzato, è stato dimostrato che un farmaco antidepressivo, la fluvoxamina, riduce la probabilità di deterioramento clinico dei pazienti affetti da COVID-19 grave [12].
È interessante notare che la fluvoxamina potrebbe facilitare l’attivazione del TMEMF e l’esposizione alla fosfatidilserina [13]. È imperativo esaminare se la fluvoxamina influisce sulla formazione di sincizi. È anche degno di valutare se l’uso combinato di farmaci anti-sincizia con altri bersagli COVID-19 produrrebbe risultati clinici migliori [14, 15].
Nel complesso, questi due documenti forniscono informazioni critiche per la comprensione di come si è verificata la sincizia durante le infezioni da SARS-CoV-2 nella struttura del virus e nei punti di vista della segnalazione cellulare e aprono nuove entrate negli studi COVID-19. Si prevede che questi nuovi risultati possano fornire informazioni per lo sviluppo di nuove strategie per combattere l’attuale pandemia.
collegamento di riferimento: https://www.nature.com/articles/s41418-021-00795-y
Discussione – link di riferimento: https://www.embopress.org/doi/abs/10.15252/embj.2021108944
La replicazione e gli effetti citopatici delle varianti SARS-CoV-2 sono oggetto di un attento esame, con risultati contrastanti in letteratura (Frampton et al., 2021; Hou et al, 2020; Leung et al, 2021; Liu et al, 2021b; Touret et al., 2021). Ad esempio, in alcuni rapporti non sono state riscontrate differenze significative nella cinetica di replicazione dei ceppi Alpha e D614G (Thorne et al, 2021; Touret et al., 2021), mentre altri hanno suggerito che Alpha potrebbe superare D614G in un test di coinfezione ( Touret et al., 2021). Altri studi hanno proposto che la mutazione N501Y possa fornire un vantaggio di replicazione, mentre altri hanno suggerito che N501Y è deleterio (Frampton et al., 2021; Hou et al., 2020; Leung et al., 2021; Liu et al., 2021b). Questi risultati discrepanti possono essere dovuti all’uso di diversi sistemi sperimentali, ceppi virali, molteplicità di infezione e tipi di cellule.
Qui, mostriamo che le varianti Alpha e Beta si replicano nella stessa misura del primo ceppo D614G in diverse linee cellulari umane e cellule delle vie aeree primarie. Inoltre, Alpha e Beta hanno indotto più fusione cellula-cellula rispetto a D614G. Un aumento della fusione è stato osservato nelle cellule U2OS-ACE2 e nelle cellule Vero naturalmente permissive. In accordo con i dati sull’infezione, la trasfezione delle proteine Alpha e Beta S, in assenza di altri fattori virali, ha prodotto sincizi significativamente maggiori rispetto a D614G, che a sua volta si è fuso più del Wuhan S.
L’analisi comparativa della microscopia video ha rivelato che Alpha S si è fusa più rapidamente, seguita da Beta, D614G e infine Wuhan. Pertanto, le varianti Alpha e Beta mostrano una maggiore formazione di sincizi mediata da S. Un limite del nostro studio risiede nel fatto che non siamo stati in grado di osservare l’espressione superficiale delle proteine S varianti in Vero e Caco2 senza perdere i grandi sincizi positivi alla proteina S.
Abbiamo quindi utilizzato le cellule 293T non fusogene per controllare l’espressione superficiale. Mostriamo inoltre che le cellule 293T che esprimono S si fondono con le cellule Vero in esperimenti di donatore/accettore. Gli esperimenti hanno confermato l’aumentata fusogenicità delle varianti in cellule con livelli simili di proteina S sulla loro superficie.
Mostriamo inoltre che Alpha e Beta rimangono sensibili alla restrizione di IFN-β1. La fusione mediata dalle rispettive proteine S è inibita dagli IFITM. Ciò estende i risultati precedenti di noi e di altri dimostrando che l’ancestrale Wuhan S è effettivamente inibito da questa famiglia di fattori di restrizione (Buchrieser et al., 2020; Shi et al., 2021).
È stato recentemente riportato in un pre-print che Alpha può portare a livelli più bassi di produzione di IFN-β1 da parte delle cellule Calu-3 infette e può essere meno sensibile al pre-trattamento di IFN-β, rispetto agli isolati virali della prima ondata (Thorne et al., 2021).
Non abbiamo rilevato qui differenze di sensibilità all’IFN-β1 tra le varianti nelle cellule Vero e U2OS-ACE2. Anche in questo caso, questi risultati discrepanti possono riflettere le differenze intrinseche tra le cellule Calu-3, Vero e U2OS-ACE2, o l’uso di diversi isolati virali.
Abbiamo quindi caratterizzato il contributo delle singole mutazioni presenti nelle proteine Alfa e Beta S alla loro rispettiva fusogenicità. L’Alpha S altamente fusogeno consiste in più mutazioni che aumentano notevolmente la fusione (P681H e D1118H) rispetto alle mutazioni che la riducono (∆69/70).
Al contrario, la variante Beta è composta da diverse mutazioni restrittive (∆242-244, K417N ed E484K) e solo una mutazione che ha aumentato leggermente la fusione (D215G). Il più forte aumento della fusione è stato provocato dalla mutazione P681H al confine S1/S2.
Questa mutazione probabilmente facilita la scissione proteolitica di S e quindi promuove la fusione cellula-cellula mediata da S. Infatti, l’analoga mutazione P681R presente nelle varianti B.1.617.2 e B.1.617.3 aumenta la scissione S1/S2 e facilita la formazione di sincizi (Ferreira et al, 2021; Jiang et al, 2020).
Da notare, un altro rapporto con valutazione indiretta della fusogenicità della variante S ha suggerito una lieve diminuzione o nessuna differenza nella fusione cellula-cellula di alfa e beta rispetto a Wuhan S (Hoffmann et al, 2021). Questi esperimenti precedenti sono stati eseguiti in cellule 293T in un secondo momento (24 ore dopo la trasfezione), il che potrebbe precludere il rilevamento di una fusione accelerata innescata dalle varianti.
Mostriamo che il legame della variante S all’ACE2 solubile è parallelo alla loro fusogenicità. Alpha ha legato il più efficientemente ad ACE2, seguito da Beta, D614G e infine Wuhan. Tuttavia, l’affinità ACE2 delle proteine S portatrici di singole mutazioni non era esattamente correlata alla fusogenicità. Ad esempio, le mutazioni N501Y e D614G hanno drasticamente aumentato l’affinità ACE2, ma solo D614G ha migliorato la fusogenicità.
La sostituzione K417N, e in misura minore ∆242-244, aveva un’affinità inferiore ad ACE2 e anche una fusione cellula-cellula limitata. La mutazione E484K limita significativamente la fusione, ma aumenta leggermente l’affinità ACE2. Ciò suggerisce che a livello delle singole mutazioni S, la relazione tra l’affinità ACE2 e l’aumento della fusogenicità non è sempre lineare. Le mutazioni varianti possono anche conferire vantaggi in modo indipendente da ACE2.
In effetti, lavori recenti hanno suggerito che la mutazione E484 può facilitare l’ingresso virale nelle cellule polmonari H522, richiedendo eparansolfati di superficie piuttosto che ACE2 (Puray-Chavez et al, 2021). Sarebbe di interesse futuro esaminare il potenziale di formazione di sincizi delle mutazioni varianti in altri tipi di cellule.
Abbiamo selezionato un pannello di 4 mAb che mostravano diversi profili di legame con le proteine Alpha, Beta, D614G e Wuhan S. Il mAb10 mirato al dominio S2 ha riconosciuto tutte le varianti ed è stato utilizzato come controllo positivo. Wuhan e D614G sono stati riconosciuti dagli altri tre anticorpi, diretti al NTD o all’RBD.
Alpha ha perso il riconoscimento dall’anti-NTD mAb71, mentre Beta non è stato riconosciuto né da mab71 né dai due anticorpi anti-RBD mAb48 e mAb 98. Esaminando il potenziale delle proteine S che trasportano singole mutazioni per legarsi agli anticorpi monoclonali umani, abbiamo scoperto che anche quelli che limitano (∆242-244, K417N) o non hanno effetto sulla fusogenicità (∆Y144) non vengono riconosciuti da alcuni mAbs.
Ciò suggerisce che le proteine varianti S hanno subito un trade off evolutivo in alcune circostanze; selezionando per le mutazioni che forniscono fuga anticorpale a scapito della fusogenicità. In accordo con i nostri risultati, l’analisi del legame della sequenza profonda e gli studi di evoluzione in vitro suggeriscono che la mutazione N501Y aumenta l’affinità con ACE2 senza disturbare la neutralizzazione dell’anticorpo (Liu et al, 2021a; Starr et al., 2021; Zahradník et al, 2021).
Le mutazioni E484K e K417N RBD nella variante Beta possono anche aumentare l’affinità ACE2, in particolare se in combinazione con N501Y (Zahradník et al., 2021) (Nelson et al, 2021). Tuttavia, il cambiamento conformazionale risultante della proteina S RBD può anche ridurre la sensibilità agli anticorpi neutralizzanti (Nelson et al., 2021).
Il lavoro futuro che valuta i cambiamenti strutturali e conformazionali nella proteina S provocati da una combinazione di singole mutazioni o delezioni può ulteriormente aiutare a chiarire l’aumentata fusogenicità e il potenziale di fuga anticorpale delle varianti.
Mentre in precedenza avevamo dimostrato che l’interazione tra la proteina S sulla membrana plasmatica con il recettore ACE2 sulle cellule vicine è sufficiente per indurre la formazione di sincizi, esistono prove convincenti dell’importanza della proteasi TMPRSS2 nell’attivazione di S (Buchrieser et al., 2020; Dittmar et al, 2021; Koch et al, 2021; Ou et al, 2021).
Abbiamo scoperto che la proteina S delle nuove varianti induceva più formazione di sincizi rispetto alle proteine D614G e Wuhan S nelle cellule Caco2 umane che esprimono ACE2 e TMPRSS2 endogeni. Tuttavia, non abbiamo rilevato alcun problema importante
differenze nell’elaborazione delle proteine S varianti da parte di TMPRSS2. Varrà la pena di caratterizzare ulteriormente come la fusogenicità delle mutazioni associate a varianti sia influenzata da altre proteasi cellulari come la furina.
La presenza di pneumociti sinciziali infetti è stata documentata nei polmoni di pazienti con COVID-19 grave (Bussani et al., 2020; Tian et al, 2020; Xu et al, 2020). La formazione di sincizi può contribuire alla replicazione e diffusione di SARS-CoV-2, all’evasione immunitaria e al danno tissutale. Un rapporto che utilizza epiteli bronchiali ricostituiti ha rilevato che l’infezione virale provoca la formazione e il rilascio di sincizi infetti che contribuiscono alla dose infettiva (Beucher et al., 2021).
La risposta anticorpale neutralizzante all’infezione da SARS-CoV-2 ha un effetto divergente sulla fusione cellula-cellula, con alcuni anticorpi che limitano la fusione mediata da S, mentre altri aumentano la formazione di sincizi (Asarnow et al, 2021). La diffusione del virus da cellula a cellula può essere meno sensibile alla neutralizzazione da parte di anticorpi monoclonali e plasma convalescente rispetto al virus acellulare (Jackson et al, 2021). È quindi possibile che le cellule sinciziali infette facilitino la diffusione virale. In questo contesto, è necessario comprendere meglio il potenziale fusogeno delle varianti SARS-CoV-2 che sono sorte e continueranno ad emergere.
Abbiamo caratterizzato qui la replicazione, la fusogenicità, il legame ACE2 e il riconoscimento anticorpale delle varianti Alfa e Beta e il ruolo delle loro mutazioni associate a S. Nonostante le intuizioni che forniamo sulla fusogenicità mediata da S delle varianti, non abbiamo affrontato i cambiamenti conformazionali che le mutazioni singolarmente o in combinazione possono suscitare.
Mostriamo inoltre che le proteine Alpha, Beta e Delta S si legano in modo più efficiente a ACE2 e sono più fusogeniche di D614G. Quali caratteristiche virologiche e immunologiche della variante Delta spieghino il suo tasso di trasmissibilità stimato più elevato rispetto all’Alpha e ad altre varianti a livello di popolazione rimane una questione in sospeso.




{kind=link}