










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Early studies indicated that monoclonal antibody treatment in immunocompromised individuals could result in within-host viral evolution preferentially affecting epitopes recognized by these antibodies, although whether this signifies a real risk of transmissible antibody resistant virus is unclear.
Emergence of SARS-CoV-2 mutations spread across multiple viral lineages capable of increased transmissibility or producing reinfection despite vaccination has caused recent concern [1, 2].
For each of these reasons, improved genomic surveillance will be critical to monitor the spread of these new variants and detect the emergence of new lineages with similarly concerning properties.
Since March 2020 we have monitored the introduction and spread of SARS-CoV-2 to the service area of the Gundersen Health System, an integrated healthcare system headquartered in La Crosse, WI and providing care in 21 counties in southwestern Wisconsin, northeastern Iowa and southeastern Minnesota.
We have performed viral whole genome sequencing on more than 1,900 positive cases from this region over the span of the pandemic. Among our major goals were monitoring for introduction of viral variants into our service area that had been recognized elsewhere by genomic surveillance measures as variants of concern. In kind, our program enabled early detection and tracking of clinically relevant polymorphisms in lineages in which they had not previously been identified elsewhere.
This suggests that convergent evolution toward some transmission-favoring phenotype may be occurring. The spike protein plays a key role in viral entry [8, 9] and is also the target of both naturally developed antibodies [10] as well as synthetic monoclonal antibodies used as treatment or post-exposure prophylaxis [11]. Studies have indicated that viruses bearing E484K are associated with a diminished response to vaccine-induced neutralizing antibodies [12].
Therapeutic monoclonal antibodies received Emergency Use Authorization (EUA) in the outpatient setting following trials demonstrating a modest reduction in the risk of hospitalization [13-15]. Initial authorization of these agents accounted for a drop in clinical utility if the prevailing circulating viral substrains shifted to include variant(s) for which these antibodies had reduced affinity, with continued availability to be based on passive monitoring of sequence data accumulating in repositories.
Epidemiologic trends subsequently lead to the withdrawal of Bamlanivimab [16] (and later Bamlanivimab/Etesevimab [17]) from widespread clinical use, though the combination was later reinstated for widespread use and expanded for use as post-exposure prophylaxis as the prevailing circulating variants changed once again [18].
Casirivimab/imdevimab became available under EUA on 11/20/20 [19] and use for post-exposure prophylaxis was subsequently authorized [20].
The potential for these treatments to select for emergence of antibody resistance mutations has been noted previously in closely monitored immunocompromised patients [21-23], but post-administration surveillance in either close contacts or the broader community has not been reported.
Between Nov 2020 and September 2021, our program administered monoclonal antibody therapies of Bamlanivimab, Bamlanivimab/Etesevimab, or Casirivimab/Imdevimab to 1,043 COVID-19 positive patients.
In parallel, our regional SARS-CoV-2 sequencing program [24-26] provided the opportunity to detect newly emerging E484K-containing lineages and ascertain the potential epidemiological association, if any, with individuals receiving monoclonal antibody therapy.
reference link : https://www.medrxiv.org/content/10.1101/2021.10.02.21264415v1
SARS-COV-2 Evolution And Impact On Recognition by Neutralizing Antibodies
The emergence of mutant SARS-CoV-2 variants and their impact on the efficacy of nAbs and vaccines has become a major concern in the current progress of the pandemic (23–25). Immune selection pressure during protracted infections are presumed to have contributed to the emergence of these variants (26).
Although as the levels of herd immunity increase through natural and vaccine induced immunity it is reasonable to speculate that this will provide greater selection pressure on the virus and we may see more mutants continue to emerge. Due to their potential to transform the strength and kinetics of the binding with ACE2, mutations that arise in the RBD are of particular interest.
The earliest variant to emerge and rapidly became dominant worldwide carried a D614G substitution and although studies suggested increased transmissibility (27–29), this variant was neutralized with existing monoclonal antibodies and convalescent sera (30–32). The emergence of the N501Y substitution was particularly concerning, which has been reported to be more transmissible than wild type and the D614G substitution (11, 33, 34).
The N501Y substitution was first seen in the B.1.1.7 variant (23) and subsequently in the B.1351 (24) and P1 variants (35). The other concerning immune escape substitution in the RBD is E484K, which is also present in the B.1.351 and P.1 variants. The B.1351 and P.1 variants also harbor K417N and K417T substitution, respectively (23–24, 35).
The presence of the N501Y substitution has been reported to increase affinity to ACE2 7-fold, and the additive combination of substitutions at 417, 484, and 501 have shown further increased affinity to ACE2 (19-fold compared to Wuhan) (11, 36). The other significant RBD substitutions are Y453F in the B.1.1.298 variant (37), L452 in the B.1.429 (25) and B.1.617 variants (38).
Recently, WHO has renamed the variants by using Greek letters to refer to the variants as: B.1.1.7 as Alpha, B.1.351 as Beta, P1 as Gamma and B.1.617.2 as Delta (39, 40) (Table 1). A variant is characterized as a variant of concern (VOC) if it demonstrates increased transmissibility, increased virulence, change in disease presentation, or reduced effectiveness of: vaccines, diagnostic testing and treatment measures (40).
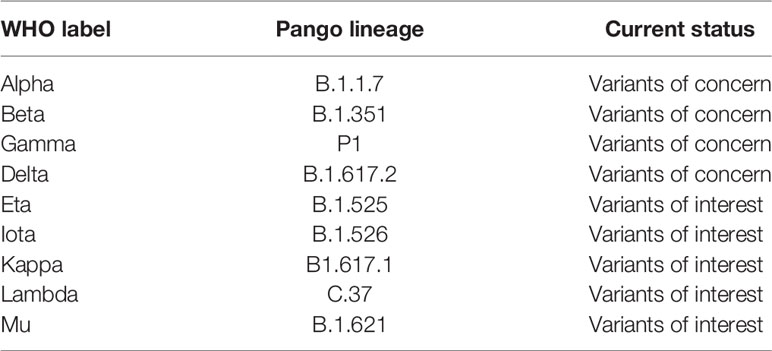
These circulating VOCs of SARS-CoV-2, including B.1.1.7 (23), B.1.351 and P.1 (35, 41) show decreased susceptibility to some SARS-CoV-2 mAbs (11, 42–44), convalescent plasma (11, 45) and sera from SARS-CoV-2 vaccinees (11, 36, 44, 46, 47). The Delta variant, B.1.617.2, has become the dominant SARS-COV-2 variant worldwide (48) and is associated with increased viral replication leading to increased transmissibility, higher viral load and severity (49, 50). The emergence of B.1.617.2 is associated with evasion of mAbs and vaccine efficacy (50, 51).
The availability of therapeutic nAbs effective against all SARS-CoV-2 variants will offer benefits for the control of the current pandemic variants and future variants, and their development therefore remains a high priority (52). The presence of broad mAbs have been described in convalescent patients and offer hope that the current vaccines might be effective (53). Most of the mAbs isolated from convalescent patients are Class 1 antibodies (18, 19).
The majority of broadly-neutralizing antibodies (bnAbs) described to-date also fall within this same class (9–11), however, bnAbs belonging to Class 4 with high potency and neutralizing breadth have been characterized (21, 22). Class 2 comprises of some potent Abs (C144, C121, COVA2-15, COVA2-37), however, their efficacy against emerging variants have not been described (19, 54). Class 3 antibodies bind to a conserved epitope and are generally unaffected by mutations (19, 55).
Understanding the different classes of broad mAbs that are generated and their persistence in convalescent recovered patients and vaccines is important for understanding the robustness of the herd immunity that is generated. Here we review some of the most promising broadly neutralizing antibodies and their mode of interaction obtained from convalescent patients. It will be important to understand how many of these broad neutralizing antibodies are retained post vaccination and natural infection.
Classification of Antibodies Based on Their Binding Mode to the RBD of the Spike Protein
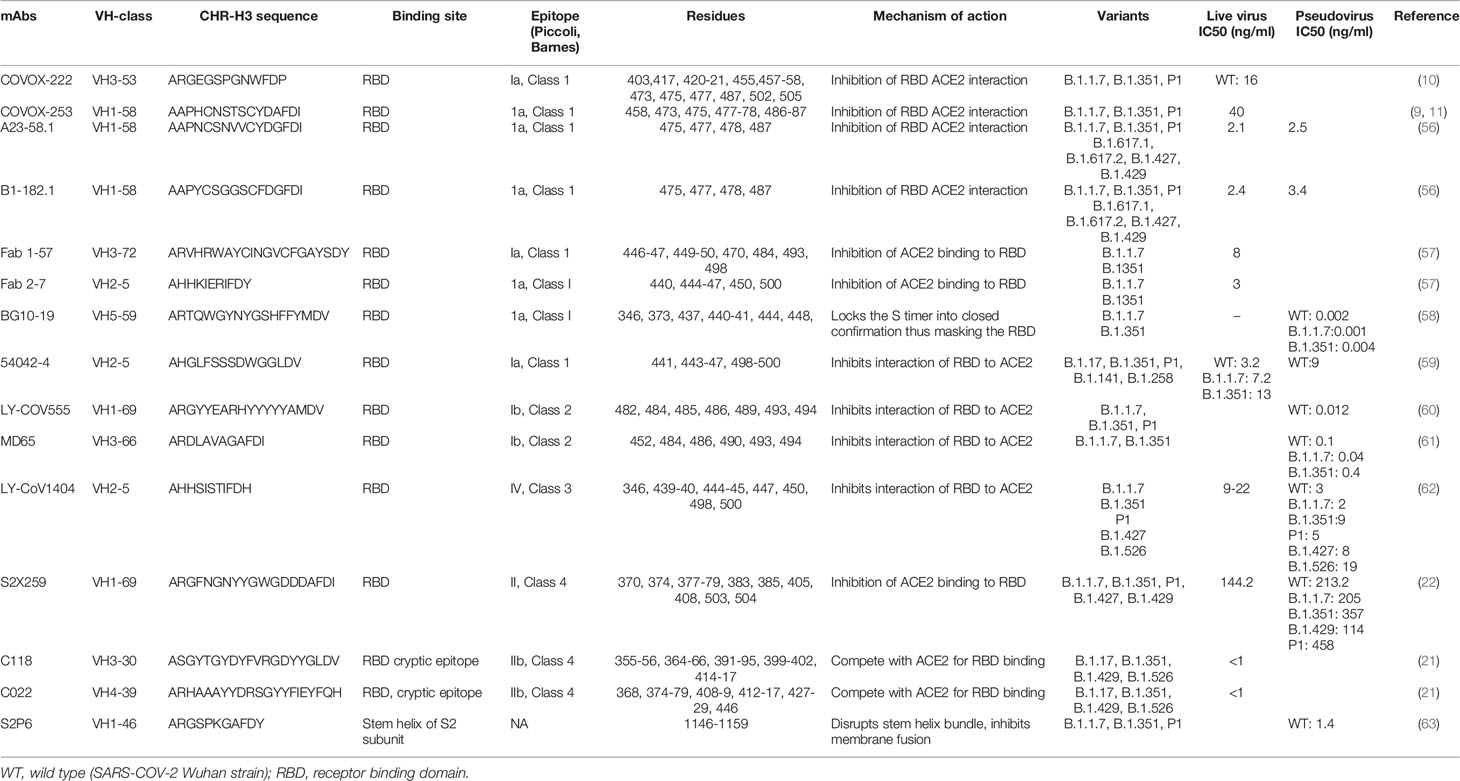
In this review, we will primarily use the Barnes classification, although the classifications by Piccoli et al. will be occasionally cross-referenced (Table 2).
Class 1 Antibodies
Class 1 Abs are the most immunodominant among RBD-targeting antibodies and are usually encoded by VH3-53 and VH3-66 germlines (5, 10, 17, 54). The family of IGHV3-53 antibodies have been described to share common binding properties and bind to a common epitope around the neck of the RBD, with an identical approach also shared by the IGHV3-66 derived Abs (5, 11, 36). The binding and neutralization of this Class of antibodies are usually abrogated by N501Y, E484K and K417N mutations (64).
Their engagement with the RBD is dictated by CDR-H1 and CDR-H2, while the CDR-H3 is short and makes few interactions (5, 10, 65). However, CDR-H3 does interact with K417 and CDR-L1 with N501, and therefore, the neutralization activity of many class I VH3–53 antibodies are compromised by the N501Y substitution in the variant viruses B.1.1.7, B.1.351, and P.1, while the added substitution at 417 in P.1 and B.1.351 has an additive negative effect on neutralizing activity (10). Hence, the neutralizing potency of the majority of the Class I mAbs are markedly reduced against the variants (66). Class 1, however, also contains several nAbs capable of neutralizing the emerging variants. In this section we discuss several of these antibodies and describe their interaction with the SARS-CoV-2 Spike.
Dejnirattisai et al. isolated the class I mAb, COVOX-222, which is derived from VH3-53, that neutralizes all three variants despite binding with two of the ACE2 binding site substitutions (10, 36) (Table 2). In the original virus, residue 417 makes a weak salt-bridge interaction with heavy-chain CDR3 residue E99 which is abolished due to a substitution to either asparagine or threonine (10).
The CDR-H3 of COVOX-222 (13 residues) is slightly longer than the majority of VH3–53 antibodies; however, this seems unlikely to be the only reason for the resilience of COVOX-222 (10, 65). There is little binding energy from the CDR3-H3, since majority of the binding energy input of the heavy chain comes from CDR-H1 and CDR-H2, which interacts weakly with RBD residue 417. Therefore many of the VH3–53 antibodies are likely to be volatile to mutations at residue 417 (K417N in B.1.351 and K417T in P1). The COVOX-22 CDR L1 interacts with residue 501 of the RBD through its P30 residue. The interaction is further strengthened by the N501Y substitution, that eventually adds to the resilience of this antibody (10). Hence, COVOX-222, a mAb of Class 1, VH3-53 gene family, despite its short CDR-H3 and binding with mutant RBD residues, is able to neutralize SARS-COV-2 variants.
Another group of class I mAbs, referred to as 55, 165, 253 and 318, have also been reported to retain nAb activity against the variants (67). These four mAbs are all IGHV1-58 class, have fewer non-silent mutations (2–5) and a longer heavy chain CDR3 (12–16 residues). mAb 55, 165 and 253 bind to the dominant neck epitope of the RBD while 318 binds to an epitope at the right shoulder. The FRNT50 titers for mAbs 55, 165, 253, and 318 are also relatively equal between Victoria strain (A.1) (67) and B.1.351, indicating that their epitopes are not impacted by the K417N, E484K and N501Y mutations (9, 11). These mAbs, despite different VH gene usage as compared to the COVOX-222, are still able to neutralize the wild type SARS-COV-2 and existing variants.
A separate group of researchers isolated two antibodies, A23-51.1 and B1-182.1, from convalescent subjects infected with the Washington-1 (WA-1) strain, which has an identical S sequence to Hu-1 (56). These antibodies showed the capacity to maintain high neutralization potency against 10 variant spike proteins including B.1.1.7, B.1.351, P1 and the highly transmissible B.1.617.2 variant.
The two antibodies, A23-58.1 and B1-182.1, share similar gene family usage in their heavy and light chains; both use IGHV1-58 heavy chains and IGKV3-20/IGKJ1 light chains and low levels of SHM. Both of these mAbs have a similar mode of binding to RBD. While binding, the RBD residue 486 dip into the crater formed by the CDRs and form a hook-like motif that is stabilized by an intra-loop disulfide bond between residues 480 and 488 while aromatic residues, including 456, 473, 486 and 489 provide 48% (299 Å2) of the epitope.
In comparison to epitopes of other antibodies, the supersite defined by common contacts of these IGHV1-58-derived antibodies had fewer interactions with residues at the mutational hotspots. The hook-like motif and CDR crater are essential for the binding within the VH1-58 public class (56). This antibody gene family combination has been proposed to be a public clonotype as it has been identified in other COVID-19 convalescent subjects (6, 9, 54).
Another study recently characterized six potently neutralizing antibodies from B cells from convalescent subjects (58). Among them, BG10-19 potently neutralized the wild-type SARS-CoV-2, B.1.1.7 (23), B.1.351 as well as the heterologous SARS-CoV pseudotyped viruses (58). BG10-19 uses five of six CDR loops to connect with a proteoglycan epitope directed up on the RBD α-1 (338–347) and α-2 (364–374) helices, with additional contacts with 436-450.
The CDRH2 and CDRH3 loops moderate the majority of RBD contacts (~760Å2), establishing strong interactions with RBD residues. CDRH1-3 and CDRL2 loops interaction establish the primary epitope recognized by BG10-19, which does not overlap with the ACE2 receptor-binding motif. Overall, BG10-19 employ a neutralization mechanism that masks engagement of the ACE2 by RBM, by locking the Spike trimer into a closed conformation (58).
Another Class I mAbs of interest are monoclonal antibodies Fab 1-57 and Fab 2-7 which bind to the RBD epitope outside the hotspot of evolutionary pressure (57). Recognition of wild-type RBD by Fab 1-57 is dominated by the heavy chain, which buries 533.7 Å surface area, with a minor 223.3 Å contribution by the light chain. With respect to the mutations in the B.1.1.7, B.1.351 and P1 variants, only residue 484 was near the binding site of Fab1-57.
Despite its proximity to the epitope, however, 484 do not interact significantly with Fab 1-57. Structural modeling of the E484K mutation showed that the K484 residue was geometrically compatible with Fab 1-57 binding at serine 29 with a hydrogen bond (57). Interaction of wild-type RBD by antibody Fab 2-7 is dominated by connection proximal to the RBD loops formed by residues 438-451 and 495-502. CDR H2 residues 54, 52, and 58 formed hydrogen bonds with RBD residues 450, the backbone amine of 445, and 447, respectively.
Light chain residue 32 also interacted with residue 440 to form a hydrogen bond. Regarding the three mutated residues, antibody Fab 2-7 bound near only N501, but the side chain of N501 pointed away from the antibody. Even though some conformational change of the 495-502 loop would occur due to N501Y mutation, this loop contributes only 225 Å2 out of 736 Å2 and contained few residues that form significant interactions with the Fab (57). The therapeutic use of Abs, such as Fab 1–57 and 2–7, which target less prevalent epitopes, could mitigate concern of mAb escape.
Along with the existing methods, newer technologies are also being deployed for this purpose (68). Kramer et al. (59) used LIBRA-seq technology (69) and identified a potent monoclonal antibody 54042-4 from a convalescent COVID-19 patient that bound and neutralized the live SARS-CoV-2 viruses, including variants of concern.
They concluded that antibody 54042-4 bound to these RBD variants (B.1.17, B.1.351, P1, B.1.141, B.1.258) at a level comparable to the binding to the RBD of Wuhan-1 isolate. The cryo-EM structure showed that 54042-4 forms a vast interface with the RBD through all three CDRs of the heavy chain, CDRL1 and CDRL3 to form a clamp over the apex of the RBM saddle.
The heavy chain forms interaction with RBD residues 443-447, while light chain interacts with residues 445 and 498-500. CDR H1 binds to 441, CDR H2 to 444 and CDR H3 to 443. CDR L1 and L3 makes hydrogen bond with 445 and 498-500. The complex interaction indicated that the spike substitutions in current VOCs are unlikely to affect the binding affinity of 54042-4.
RBD residue N501 lies outside of the 54042-4 epitope, while the Cα atoms of E484 and K452 are 18 and 14 Å far from the nearest 54042-4 residue, respectively. The use of modern technology like LIBRA-seq enables high-throughput concurrent determination of B cell receptor sequence and antigen reactivity at the single-cell level, facilitating the candidate selection and characterization process, and also expedite the development of broadly-neutralizing antibodies.
Class 1 is comprised of a diverse array of mAbs with strong neutralization breadth and potency. Despite variation in their VH gene usage, all the above mAbs can effectively neutralize the emerging variants of SARS-COV-2. These antibodies could be used alone or in combination with other classes of antibodies in manufacturing therapeutic antibodies and also guide the formulation of next generation vaccines.
Class 2 Antibodies
Class 2 comprises of some potent Abs (C144, C121, COVA2-15, COVA2-37) that can bind to the RBD in both the up and down confirmation. In general, through their efficacy against emerging variants have not been described (19, 54). However, two Class 2 mAbs and the impact of known mutations have been well described. LY-CoV555 (clinical name bamlanivimab), currently used in the clinical setting in combination with etesivimab, is a potent anti-spike neutralizing antibody (70, 71).
Structural analysis revealed that it binds both in up and down confirmation of the RBD, and the binding of this mAb to RBD is not affected by the N501Y mutation; however, the E484K mutation abolishes binding (60). Against B.1.351, activities of bamlanivimab and another therapeutic mAb REGN10933 belonging to Class 1 antibodies are also abolished (66). Another Class 2 antibody, MD65, has a binding pattern similar to LY-CoV555 but unlike LY-CoV555, the E484K mutation did not affect its binding efficacy (61).
The in vivo assessment on K18-hACE2 transgenic mice suggested that MD65 exhibited efficacy against B.1.1.7, B.1.351 and P1 variant (61). So, it appears that broadly reactive Class 2 mAbs can be developed despite the shortcomings of bamlanivimab against B.1.351.
Class 3 Antibodies
Antibodies targeting the Class 3 epitope can bind with the RBD in both “up” and “down” states (18, 19). The class 3 epitope is highly conserved in Sarbecovirus clades 1, 2, and 3, indicating it is a good target for broad neutralizing antibodies and suggests it is functional conserved and less likely to be associated with immune (55, 72). The Class 3 antibodies bind outside of the ACE-2 binding region and hence, provide the potential for synergistic effects when combined with nAbs that intercept ACE2 binding.
The Class 3 monoclonal antibody LY-CoV1404 was isolated from a high-throughput screen of peripheral blood mononuclear cells obtained from a convalescent subject 60 days after symptom onset (62). Using authentic and pseudoviruses neutralization assays, researchers showed that LY-CoV1404 maintains potent neutralizing activity against multiple variants including B.1.1.7, B.1.351, B.1.427, P.1 and B.1.526. Interestingly, LY-CoV1404 share 92% amino acid sequence identity in the variable regions of both its heavy and light chains, to antibody Fab 2-7 (57), though they were discovered independently from different patients suggesting it maybe a publicly shared repertoire (62).
Both antibodies also append to the RBD in a similar way. LY-CoV1404 binds to a region overlapping the ACE2-interacting site of the spike that is accessible in open and closed state of the RBD. While this property would advise that this is a Class 2 antibody (5, 18), the location of the epitope is identical to mAb S309, a class 3 binder (55). Although LY-CoV1404 binding epitope includes residues N501 and N439, they can bind the B.1.1.7 and B.1.351 variants and are neutralized as strongly as wild type virus (62).
The above findings suggest that LY-CoV1404 represents a potent mAb with broad neutralizing range, possess the property of both Class 2 and 3 antibodies, has a relatively conserved epitope and could well be deployed to address the concern of emerging variants.
LY-CoV1404’s potent neutralization of SARS-CoV-2 allows for exploration of lower clinical doses, which may support subcutaneous administration and has the potential to provide a long-term complement to vaccines in the likely event that COVID-19 becomes endemic. Class 3 nAbs supplement the anti-SARS-CoV-2 antibody repertoire and can be effectively utilized in therapeutic combinations with class 1 or 2 nAbs (18).
Another group of antibodies in class 3 are VIR-7831 and VIR-7832; dual action mAbs derived from the parent antibody S309–an antibody obtained from a SARS-CoV survivor (55). These mAbs have been engineered to have an extended half-life and improved lung bioavailability (73) and target to an epitope located around residue N343 that is highly conserved among the Sarbecovirus, and neutralize live and pseudotyped virus against B.1.1.7, B.1351 and P1 variants. They also exhibit potent effector function and confer antibody dependent cellular cytotoxicity and antibody dependent cellular phagocytosis in vitro (74). The mAb VIR-7831 has been provided emergency use authorizations (EUA) by the US FDA for treatment of mild to moderate COVID-19 (75).
Class 4 Antibodies
Class 4 Abs bind to the highly conserved, cryptic epitope on the RBD outside the RBM (18). The majority of mAbs described previously are cross-reactive but weakly neutralizing (1, 17, 76–78). However, Jette et al. (21) characterized two Class 4 anti-RBD antibodies, C118 and C022, that were obtained from COVID-19 donors and revealed broad recognition and potent neutralization of SARS-CoV-2 variants (21).
They found that C118 and C022 Abs neutralized four SARS-CoV-2 variants (B.1.17, B.1.351, B.1.429, B.1.526). The structure analysis showed that both mAbs recognized an epitope that is highly conserved at the base of the RBD, which is disclosed only in ‘up’ conformations. C118 and C022 use four of six CDR loops to bind to the epitope that stretches towards the RBD ridge close to the ACE2 binding region, and C022 includes an additional overlapping interacting residue at 417.
CDRH3 and CDRL2 loops, and portions of framework region L3 of both antibodies dominate the RBD contacts and develop polar and van der Waals interactions that accounts for 71% of the epitope buried surface area (21). C022 and C118 form considerable backbone interactions with RBD, with 9 and 10 H-bonds formed with the RBD, respectively contributing to their cross-neutralization across variants and breadth, as these interactions would mediate binding despite side chain substitutions.
The isolation and characterization of nAbs targeting conserved RBD epitopes that possess dual advantage of breadth across Sarbecoviruses and higher resistance to neutralization escape is necessary to fight this pandemic (79). Due to the conservation at the Class IV epitope, antibodies targeting this site are of interest. S2X259, is another class 3 antibody isolated (22) from a convalescent patient which broadly neutralizes entry of SARS-CoV-2 including the B.1.1.7, B.1.351, P.1 and B.1.427/B.1.429 variants. This antibody acts through inhibition of ACE2 binding to the RBD.
They also performed several experiments to show that this antibody is effective against a wide spectrum of human and zoonotic sarbecoviruses and retain a high barrier to the emergence of resistant mutants (22). S2X259 targets a glycan-free, cryptic epitope within antigenic site IIa and binds with the RBD using both heavy and light chains contributing two thirds and one third of the paratope surface buried upon binding, respectively (1, 22).
S2X259 uses CDR H1-H3, L1 and L3 to interact with residues 369-386 forming two α-helices and an intervening β-sheet. The S2X259 epitope is highly conserved in SARS-CoV-2 viruses and does not contain prevalent mutant residues, such as L452R, S477N or N439K. The mAb binds with the backbone of residue N501 and not its side chain, circumvents the residues 417 and 484 which could explain the preserved neutralizing potency against B.1.1.7, P1, B.1.351 and B.1.429 (22).
Due to its conserved and cryptic epitope, Class 4 antibodies are better assets for neutralization across multiple variants and thereby potentially protect against emergent Sarbecoviruses (21, 22).
reference link : https://www.frontiersin.org/articles/10.3389/fimmu.2021.752003/full?fbclid=IwAR23Dbry666btcHAdEsk0kMrI-XejO6XXsX44DkNEa2A-DF5Nkqh91Y88aE



{kind=link}