









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Let’s start with representing the situation today…
Variants of Concern (VOC)
| WHO label | Lineage + additional mutations | Country first detected (community) | Spike mutations of interest | Year and month first detected | Impact on transmissibility | Impact on immunity | Impact on severity | Transmission in EU/EEA |
|---|---|---|---|---|---|---|---|---|
| Beta | B.1.351 | South Africa | K417N, E484K, N501Y, D614G, A701V | September 2020 | Increased (v) (1) | Increased (v) (2, 3) | Increased (v) (4, 5) | Community |
| Gamma | P.1 | Brazil | K417T, E484K, N501Y, D614G, H655Y | December 2020 | Increased (v) (6) | Increased (v) (7) | Increased (v) (5) | Community |
| Delta | B.1.617.2 | India | L452R, T478K, D614G, P681R | December 2020 | Increased (v) (8) | Increased (v) (9-11) | Increased (v) (10, 12) | Community |
| Omicron | B.1.1.529 | South Africa and Botswana | (x) | November 2021 | Unclear (v) (13-15) a | Increased (v) (16) | Reduced (v) (17-23) b | Dominant |
x: A67V, Δ69-70, T95I, G142D, Δ143-145, N211I, Δ212, ins215EPE, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H, N764K, D796Y, N856K, Q954H, N969K, L981F
a: The observed increased growth rate may be due to increased inherent biological transmissibility, contextual factors such as transmitting in population groups with increased contact rates, or escape from immunity which increases the size of the susceptible population.
b: Preliminary studies show reduced risk of hospitalisation, but more data from EU/EEA countries is required to determine if this effect is observed across population groups (e.g. by age, vaccination and prior infection status). Conclusive evidence on mortality risk is not yet available.
All sub-lineages of the listed lineages are also included in the variant, e.g., C.37.1 is included in Lambda as it is a sub-lineage of C.37.
Variants of Interest (VOI)
| WHO label | Lineage + additional mutations | Country first detected (community) | Spike mutations of interest | Year and month first detected | Impact on transmissibility | Impact on immunity | Impact on severity | Transmission in EU/EEA |
|---|---|---|---|---|---|---|---|---|
| Mu | B.1.621 | Colombia | R346K, E484K, N501Y, D614G, P681H | January 2021 | Increased (m) (24) | Increased (m) (25) | No evidence | Sporadic/Travel |
| Lambda | C.37 | Peru | L452Q, F490S, D614G | December 2020 | No evidence | Increased (v) (26, 27) | No evidence | Sporadic/Travel |
| n/a | AY.4.2 | United Kingdom | L452R, T478K, D614G, P681R, A222V, Y145H | June 2021 | Increased (v) (28) | Similar (v) (28, 29) | Similar (v) (28) | Community |
n/a: not applicable, no WHO label has been assigned to this variant at this time
All sub-lineages of the listed lineages are also included in the variant, e.g., AZ.1 is included in B.1.1.318 as it is a sub-lineage of it.
Variants under monitoring
These additional variants of SARS-CoV-2 have been detected as signals through epidemic intelligence, rules-based genomic variant screening, or preliminary scientific evidence. There is some indication that they could have properties similar to those of a VOC, but the evidence is weak or has not yet been assessed by ECDC. Variants listed here must be present in at least one outbreak, detected in a community within the EU/EEA, or there must be evidence that there is community transmission of the variant elsewhere in the world.
| WHO label | Lineage + additional mutations | Country first detected (community) | Spike mutations of interest | Year and month first detected | Impact on transmissibility | Impact on immunity | Impact on severity | Transmission in EU/EEA |
|---|---|---|---|---|---|---|---|---|
| n/a | B.1.1.318 | Unclear (b) | E484K, D614G, P681H | January 2021 | No evidence | Increased (m) (25) | No evidence | Detected (a) |
| n/a | B.1.617.2 + K417N | United Kingdom | L452R, T478K, D614G, P681R, K417N | June 2021 | No evidence | No evidence | No evidence | Detected (a) |
| n/a | C.1.2 | South Africa | D614G, E484K, H655Y, N501Y, N679K, Y449H | June 2021 | Increased (m) (24) | Increased (m) (25) | No evidence | Detected (a) |
| n/a | B.1.617.2 + E484X (d) | India | L452R, T478K, D614G, P681R, E484X (d) | April 2021 | No evidence | No evidence | No evidence | Detected (a) |
| n/a | B.1.617.2 + Q613H | India | L452R, T478K, D614G, P681R, Q613H | April 2021 | No evidence | No evidence | No evidence | Detected (a) |
| n/a | B.1.617.2 + Q677H | India | L452R, T478K, D614G, P681R, Q677H | April 2021 | No evidence | No evidence | No evidence | Detected (a) |
| n/a | B.1.640 | The Republic of Congo | D614G, F490R, N394S, N501Y, P681H, R346S, Y449N, 137−145del | September 2021 | No evidence | No evidence | No evidence | Detected (a) |
n/a: not applicable, no WHO label has been assigned to this variant at this time
All sub-lineages of the listed lineages are also included in the variant, e.g., AZ.1 is included in B.1.1.318 as it is a sub-lineage of it.
De-escalated variants
| WHO label | Lineage + additional mutations | Country first detected (community) | Spike mutations of interest | Year and month first detected | Impact on transmissibility | Impact on immunity | Impact on severity | Rationale for de-escalation | |
|---|---|---|---|---|---|---|---|---|---|
| Alpha | B.1.1.7 | United Kingdom | N501Y, D614G, P681H | September 2020 | Increased (v) (24) | Similar | Increased (v) (5, 30) | Drastically reduced circulation in the EU/EEA following the emergence of Delta; little evidence of impact on vaccine induced immunity | |
| n/a | B.1.1.7+E484K | United Kingdom | E484K, N501Y, D614G, P681H | December 2020 | Increased (v) (24) | Increased (v) (25, 31) | Increased (v) (30) | Very low levels of circulation in the EU/EEA | |
| Epsilon | B.1.427/B.1.429 | USA | L452R, D614G | September 2020 | Unclear (32) | Increased (v) (32) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA and available data indicating that vaccines and treatments are effective against such variant | |
| n/a | B.1.616(c) | France | V483A, D614G, H655Y, G669S | February 2021 | Detection (c) (33) | No evidence | No evidence | Not detected since 2021-04-23 (34) | |
| Eta | B.1.525 | Nigeria | E484K, D614G, Q677H | December 2020 | No evidence | Increased (m) (25, 35) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| Theta | P.3 | The Philippines | E484K, N501Y, D614G, P681H | January 2021 | Increased (m) (24) | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| Kappa | B.1.617.1 | India | L452R, E484Q, D614G, P681R | December 2020 | Increased (v) (36) | Increased (v) (37-40) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.620 | Unclear (b) | S477N, E484K, D614G, P681H | February 2021 | No evidence | Increased (m) (25, 41) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.617.3 | India | L452R, E484Q, D614G, P681R | February 2021 | Increased (m) [1] | Increased (m) (25, 32) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.214.2 | Unclear2 | Q414K, N450K, ins214TDR, D614G | December 2020 | No evidence | No evidence | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | A.23.1+E484K | United Kingdom | V367F, E484K, Q613H | December 2020 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | A.27 | Unclear (b) | L452R, N501Y, A653V, H655Y | December 2020 | Increased (m) (24) | Increased (m) (32) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | A.28 | Unclear (b) | E484K, N501T, H655Y | December 2020 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | C.16 | Unclear (b) | L452R, D614G | October 2020 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.351+P384L | South Africa | P384L, K417N, E484K, N501Y, D614G, A701V | December 2020 | Increased (v) (1) | Increased (v) (2, 3) | Unclear (4) | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.351+E516Q | Unclear (b) | K417N, E484K, N501Y, E516Q, D614G, A701V | January 2021 | Increased (v) (1) | Increased (v) (2, 3) | Unclear (4) | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.1.7+L452R | United Kingdom | L452R, N501Y, D614G, P681H | January 2021 | Increased (v) (24) | Increased (m) (32) | Increased (v) (30) | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.1.7+S494P | United Kingdom | S494P, N501Y, D614G, P681H | January 2021 | Increased (v) (24) | Increased (m) (42) | Increased (v) (30) | No longer detected or detected at extremely low levels in the EU/EEA | |
| Iota | B.1.526 | USA | E484K, D614G, A701V | December 2020 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.526.1 | USA | L452R, D614G | October 2020 | No evidence | Increased (m) (32) | No evidence | Lineage withdrawn from Pango | |
| n/a | B.1.526.2 | USA | S477N, D614G | December 2020 | No evidence | No evidence | No evidence | Lineage withdrawn from Pango | |
| Zeta | P.2 | Brazil | E484K, D614G | January 2021 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | B.1.1.519 | Mexico | T478K, D614G | November 2020 | No evidence | Increased (m) (32) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | AV.1 | United Kingdom | N439K, E484K, D614G, P681H | March 2021 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | AT.1 | Russia | E484K, D614G, N679K, ins679GIAL | January 2021 | No evidence | Increased (m) (25) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | C.36+L452R | Egypt | L452R, D614G, Q677H | December 2020 | No evidence | Increased (m) (32) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA | |
| n/a | P.1+P681H | Italy | D614G, E484K, H655Y, K417T, N501Y, P681H | February 2021 | No evidence | Unclear (43, 44) | No evidence | No longer detected or detected at extremely low levels in the EU/EEA |
n/a: not applicable, no WHO label has been assigned to this variant at this time
All sub-lineages of the listed lineages are also included in the variant, e.g., B.1.429.1 is included in B.1.427/B.1.429 as it is a sub-lineage of B.1.429.
(a) No assessment of transmission is given for variants in the monitoring category, only detected/not detected.
(b) The earliest detections from several different countries are close in time and there is no clearly demonstrated travel link to a specific country that explains the detections.
(c) The property of concern for this variant was the fact that there are reports of difficulties associated with detecting it in upper respiratory tract samples. These difficulties were not caused by primer-template mismatch but rather by the virus not being present in sufficient quantities in the upper respiratory tract.
(d) Any amino acid substitution
. . .
SARS-CoV-2, like other RNA viruses, undergoes genome modifications as a result of virus replication. Although the viral RNA-dependent RNA polymerase (RdRP) complex has proofreading ability that makes nucleotide substitutions less likely than in other RNA viruses, it should be noted that the proofreading mechanism does not prevent deletions in the viral genome.
Furthermore, although coronaviruses have a proofreading apparatus, their genomes are still susceptible to recombination and other copy-choice transcriptional errors [9]. These characteristics, together with the high prevalence of the virus in the human population across broad geographical regions, have led to the emergence of new virus variants that differ from the original virus circulating in China since December 2019 in terms of transmission, pathogenicity, diagnostics, and vaccine effectiveness. With coronaviruses, variants are common; however, a variant becomes a VOC when its variations have clinical or public-health consequences [10].
In addition to VOCs, the CDC has introduced a different classification for the new variants, which includes three groups: VOIs, VOCs, and Variants of High Consequence (VOHCs) [11]. Based on the CDC’s classification, a variant will be declared a VOC when it has some properties such as widespread interference with diagnostic test targets, reduced susceptibility to one or more therapeutic classes, decreased neutralization by antibodies elicited during previous infection or vaccination, reduced vaccine-induced protection from severe disease, or increased transmissibility and disease severity [11].
The three global VOCs are B.1.1.7 (Alpha), B.1.351 (Beta), and P.1 (Gamma), which originally emerged in the United Kingdom, South Africa, and Brazil, respectively [12,13,14]. B.1.617.2 (Delta), a variant of interest first detected in India last year has recently been declared a variant of global concern by WHO, with preliminary research finding that it spreads more rapidly than previous variants. This fourth variant was classified as a global threat on May 10, 2021 (Supplementary Table S1) [15].
Other variants of interest, such as B.1.427/B.1.429 in the United States, have become prevalent and predominant in particular regions and countries and are anticipated to become VOCs in the future [16]. The US CDC has classified the variants B.1.427 and B.1.429 as VOCs, although WHO has not yet classified them as such so far.
B.1.1.7 variant (Alpha)
In mid-September 2020, the first case was found in London and Kent, United Kingdom, and in December 2020, 1108 cases of infection with this variant had been identified in the UK. As of March 16, 2021, the Alpha variant had become dominant in 21 countries: the United Kingdom, Ireland, Bulgaria, Slovakia, Israel, Luxembourg, Portugal, Denmark, the Netherlands, Norway, Italy, Belgium, France, Austria, Switzerland, Liechtenstein, Germany, Sweden, Spain, Malta, and Poland. In the United States, the Alpha variant first appeared in November 2020, and the number of cases grew from 76 cases in 12 states on January 13 to 7501 cases in all 50 states by March 23 [17].
On Jan 22, 2021, the UK Government’s New and Emerging Respiratory Virus Threats Advisory Group (NERVTAG) published that patients infected with the Alpha variant showed no significant difference in their risk of hospitalization or death (risk ratio [RR] 1.00 [95% CI 0·58–1.73]). However, the next cohort study with additional data showed an RR of 1.65 (1.21–2.25). Additionally, the London School of Hygiene & Tropical Medicine and Imperial College London reported a relative hazard of death of 1.35 (95% CI 1.08–1.68) and a case fatality rate of 1.35 (1.18–1.56) [18]. This strain has a significantly higher rate of transmission (R0) and may be higher among children, while older people appear to be less affected [19].
Graham and colleagues found that individuals infected with the Alpha variant had more variation in their symptoms than previous COVID-19 patients and that loss of taste and smell was less common among patients with the Alpha variant, whereas cough, sore throat, myalgia, and fatigue were more frequent [20, 21].
Examination of S gene mutations showed that the mutation N501Y in the receptor-binding domain (RBD) alters the affinity of virus binding to ACE2, that the Δ69-70 deletion in the spike contributes to evasion of the immune response, and that P681H affects the furin cleavage site, altering the susceptibility of the spike to furin cleavage and the infectivity of the virus [22]. Mutations such as N501Y in the receptor-binding domain of the spike protein possibly affect transmissibility, pathogenicity, and vaccine efficacy [22]. A large number of non-synonymous substitutions have been reported for this variant.
A deletion at positions 69 and 70 of the spike protein (Δ69–70), which is associated with diagnostic real-time PCR failure (S gene target failure: SGTF), has been reported to be a marker for the Alpha variant, which was given the name VOC 202012/01 by Public Health England (PHE) [23, 24].
On 2 February 2021, Public Health England had detected a number of Alpha VOC-202012/01 genomes with the E484K mutation, which was called Variant of Concern 202102/02 (VOC-202102/02) [25].
N501Y, a change from asparagine (N) to tyrosine (Y) in the RBD, can increase ACE2 binding affinity and lead to the virus becoming more infectious [26]. Position 501 is in the interior of the spike protein and affects the efficacy of neutralization of virus, but there are currently no neutralization data on N501Y available using polyclonal sera from natural infections. The 69–70 deletion has eluded the immune response in some immunocompromised patients and is associated with other changes in the RBD.
Other mutations, including T1001I, A1708D, I2230T, and del3675–3677SGF in the ORF1ab gene, del69–70HV, del144Y, N501Y, A570D, D614G, P681H, T761I, S982A, and D1118H in the S gene, Q27stop, R52I, and Y73C in ORF8, and D3L and S235F in the N gene have been reported, but their effect is not known (Fig. 1, Tables 1, 2) [27].
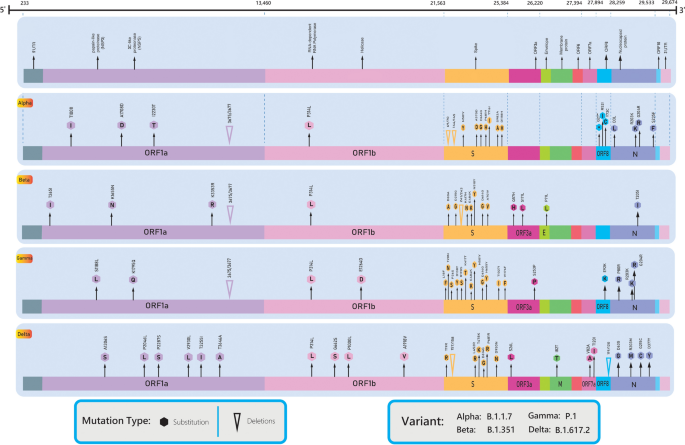
Table 1 Full list of nomenclature, origin, and mutations in the spike gene of VOCs/VOIs
From: The ins and outs of SARS-CoV-2 variants of concern (VOCs)
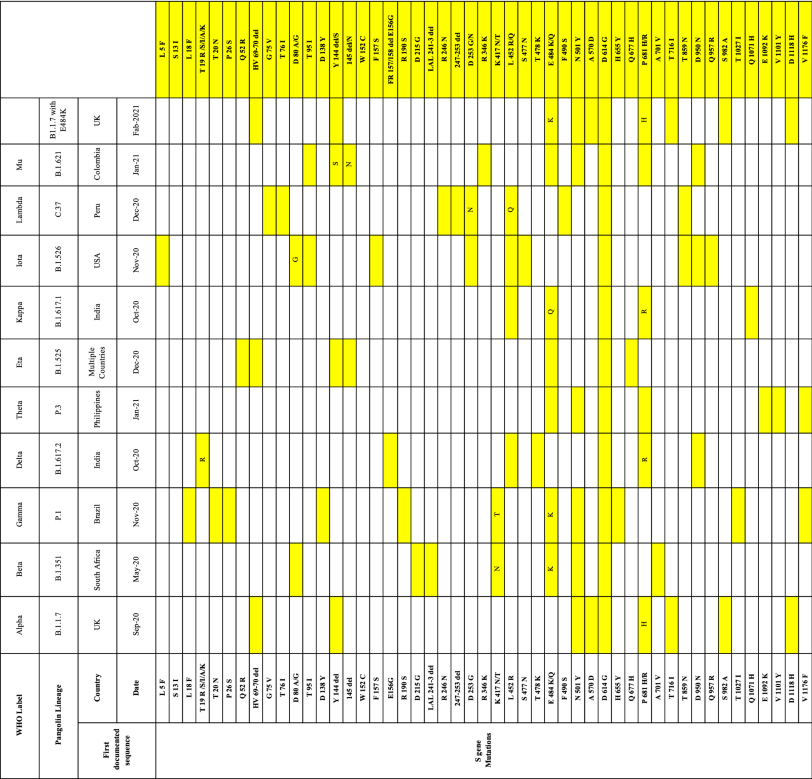
Table 2 Full list of mutations in the S gene and other genes of VOCs
From: The ins and outs of SARS-CoV-2 variants of concern (VOCs)

The ChAdOx1 nCoV-19 vaccine (developed by the University of Oxford and AstraZeneca) showed approximately 75% efficacy against the Alpha variant, compared with 84% against other lineages [20].
The ORF8 protein is an immunoglobulin-like protein that modulates pathogenesis, mediates degradation of major histocompatibility complex I (MHC-I), and suppresses the type I interferon (IFN)-mediated antiviral response. Genetic sequencing of VOC-202012/01 has shown the presence of a Q27stop mutation that inactivates the ORF8 protein, and this mutation is associated with symptoms and disease outcome [28, 29].
B.1.351 variant (Beta)
In early October 2020, the Beta variant emerged in Nelson Mandela Bay (a metropolitan area of the Eastern Cape province of South Africa). The new variant, named 501Y.V2, is also known as the B.1.351 lineage, or 20H/501Y.V2 Next strain clade. By December 2020, it had spread to Botswana, France, Scotland, South Korea, Sweden, Switzerland, and the UK [30].
The Beta variant is 50% more transmissible than the variants that were circulating previously in South Africa, and this resulted in a change in disease severity. Beta variant mutations did not impact the sensitivity of a real-time reverse transcription polymerase chain reaction (RT-PCR) assay targeting the RdRp, Nsp2, E, and N genes [31].
The potential for reinfection with the Beta variant is relatively high because mutations in the Beta variant allow escape from neutralizing antibodies in COVID-19 convalescent plasma [32].
The Beta variant contains nine spike mutations: L18F, D80A, D614G, 242–244del, and R246I in the ??N-terminal domain?? (NTD), K417N, E484K, and N501Y in the RBD, and A701V near the S2 subunit furin cleavage site (Fig. 1, Table 2) [33].
Three of the mutations in the spike region, K417N, E484K, and N501Y, are more important than the other five. All three are in the RBD and help the virus to attach more easily to human cells and increase the binding affinity for the ACE2 receptor [34].
P.1 variant (Gamma)
Lineage P.1, also known as 20J/501Y.V3, Variant of Concern 202101/02 (VOC-202101/02), or the Brazilian variant, was first detected on 6 January 2021 by the National Institute of Infectious Diseases (NIID), Japan, in people who had arrived in Tokyo having visited Amazonas, Brazil. It caused widespread infection in the city of Manaus [35]. The Gamma variant caused higher viral loads and showed 1.4–2.2 times higher transmissibility than earlier strains, especially in young patients (no gender differential) [36].
The Brazil–United Kingdom CADDE Centre on 12 January 2021 confirmed the presence of the Gamma variant in Manaus, Amazonas State [35]. Two distinct sub-variants of the Gamma variant, 28-AM-1 and 28-AM-2, carry the K417T, E484K, N501Y mutations, and both originated in Brazil [37].
The Gamma variant contains 10 mutations in the spike (L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, H655Y, and T1027I) in addition to D614G. K417T, E484K, and N501Y are in the RBD; L18F, T20N, P26S, D138Y, and R190S are in the NTD; and H655Y is near the furin cleavage site (Fig. 1, Table 2) [38, 39]. The K417T and N501Y mutations are present in both the Alpha and Beta variants and help the virus to attach tightly to human cells. E484K can help the virus evade antibodies. A deletion in the orf1b protein (del11288-11296 [3675-3677 SGF]) has been reported [37].
The Gamma variant is different from the other Brazilian lineage P.2 (‘B.1.1.28.2’ or ‘VUI-202101/01’). P.2 only has the E484K mutation in the RBD but neither the N501Y nor the K417T mutation [40].
The novel P.2 lineage was first detected by genome sequencing in October 2020, and during October and November 2020, the E484K mutation was found in all P.2 samples [41]. The P.2-specific mutations are E484K in the S gene, A119S in the N gene, 5’UTR C100U, and L3468V and synC11824U in the ORF1ab gene. Other mutations include 3’UTR C29754U, F120F (synC28253U) in ORF8, M234I in the N gene, and L3930F and synA12964G in ORF1ab [14, 42].
B.1.617.2 variant (Delta)
On May 10, 2021, WHO declared the B.1.617 lineage, originally identified in India, the fourth global VOC. This lineage has three sublineages: B.1.617.1, B.1.617.2, and B.1.617.3. There are two signature mutations for the Delta variant in the spike protein, at position 452 in the RBD region and position 681 in the furin cleavage site between S1 and S2, together with the primary D614G mutation.
The Delta variant is an exception in the B.1.617 lineage and does not have a substitution at residue 484 (Fig. 1, Table 2) [43, 44]. On June 1, 2021, WHO reported that only the B.1.617.2 sublineage, which was designated the Delta variant, remained a VOC, and sublineage B.1.617.1 was designated the Kappa variant and was classified as a VOI. B.1.617.3 was neither labeled nor reclassified [45].
In comparison to the first wave, other reports indicate that case numbers grew more quickly during the most recent surge when Alpha and Delta variants were circulating (February 11, 2021). Preliminary data and estimates revealed very high test positivity rates (TPRs) and rising case fatality rates (CFRs) in the second surge of COVID-19, with Alpha and Delta variants circulating in India [46]. The Delta variant affects serological neutralization and transmissibility, as explained below.
There is a Delta plus variant, which has an additional K417N mutation that allows the virus to better attack lung cells and potentially escape vaccine protection. The Delta plus variant is rapidly spreading in India, the UK, and other countries [47].
Features of VOCs
The emergence of a VOC indicates that increased spread or pathogenicity of the virus and increased mortality due to COVID-19 must be seriously considered. Mutations accumulate in the viral genome as a result of free circulation of SARS-CoV-2 in communities, which could influence viral transmission, mortality, detection, and immune escape [48].
Transmissibility
The accumulation of silent mutations in the SARS-CoV-2 genome provided a selective advantage in which the Alpha variant rapidly replaced common non-VOCs during autumn 2020 in the UK. Remarkably, as of 27 July 2021, this variant has been reported in 182 countries (Fig. 2) [1, 48,49,50].
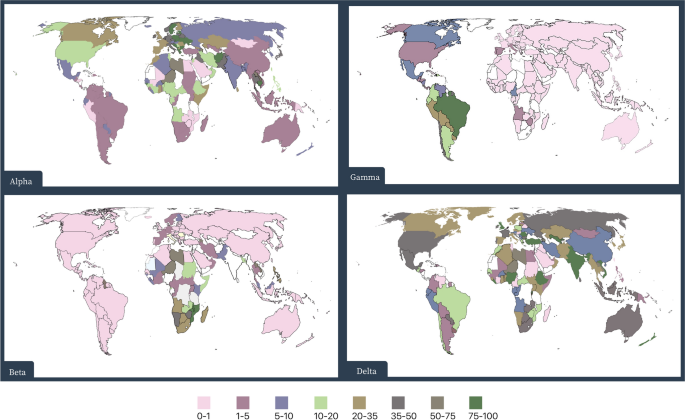
Before the spread of the Delta variant in the UK, SGTF data showed that the Alpha variant, due to increased viral loads, had higher transmissibility than non-VOC lineages in all regions of England and that it was the dominant strain [48, 51].
Using multiple models, the Alpha variant in the UK was found to have a higher reproduction number, R, than other strains. Similar increases were reported in Switzerland, Denmark, and the United States [48, 52]. The transmissibility of the Alpha variant has been estimated in various modeling studies. Rates of 68%, 75%, 56%, 40%, and 36% were reported by Vöhringer et al., Leung et al., Davies et al., Tuite et al., and a study from Denmark, respectively [49, 53, 54].
Davies et al. suggested a relationship between the Alpha variant and more-severe illness compared to previous circulating SARS-CoV-2 variants [54]. After adjustment for confounding factors, Frampton and colleagues found that patients with Alpha and non-Alpha infections showed similar disease severity and clinical outcomes [51]. Some researchers have suggested that the increased transmissibility of the variant is related to increased stability in the environment.
However, Schuit and colleagues reported that the stability of the Alpha variant in aerosols is similar to that of the earlier isolates, and therefore, the increased transmissibility of the Alpha variant is not associated with increased virus survival in the environment [55]. Spike mutations, including N501Y, P681H, and 69–70del, are key factors in the increased transmissibility of the B.1.1.7 variant. N501Y, which also is present in the Beta and Gamma variants, increases the affinity of the RBD for ACE2.
Experimental studies have shown that N501Y increases the rate of virus replication in both cell culture (human upper airway cells) and animal models (the upper respiratory tracts of hamsters) [56]. The P681H mutation, which is close to the S1/S2 furin cleavage site, increases transmissibility by increasing the efficiency of S1/S2 cleavage in human airway epithelial cells [57, 58].
The rapid transmission observed with the Gamma variant results in a higher estimated R value for this lineage than for other common circulating lineages (non-VOCs), and this is associated with higher viral loads [59]. In a comparison of Gamma variant infections with non-Gamma infections, Naveca et al. showed that Gamma variant infections were significantly associated with lower Ct values, ~ 10-fold higher viral loads, and equal levels of infectivity for individuals >18 years old [60].
Phylodynamic modeling revealed that the Gamma variant could be 2.2-2.5 times more transmissible than the circulating parental lineage, with a probability of reinfection of 6.4% (CI 5.7–7.1%) and a 25-61% higher probability of evading protective immunity. By 27 July 2021, this variant had been reported in 81 countries (Fig. 2) [1, 50, 59, 61, 62].
Preliminary results suggest that the Beta variant, with several mutations in the RBD site of the S gene (including E484K and N501Y), is probably 50% more transmissible and capable of immune escape than a co-circulating virus in South Africa. As of 27 July 2021, 131 countries have reported detection of this variant (Fig. 2) [1, 49,50,51].
Increased transmissibility and immune escape give the 501Y.V2 variant a selective advantage to outcompete other circulating SARS-CoV-2 variants by both rapid expansion and possible reinfection. Substitutions in the RBD of the spike protein (E484K, K417N, and N501Y) in the 501Y.V2 variant are associated with enhanced binding affinity to human ACE2, which may increase viral transmissibility. As a result of the selective advantage of 501Y.V2, the other circulating lineages in South Africa are rapidly being replaced by this variant [30]. In addition to N501Y, the Gamma and Beta variants have mutations that enhance transmissibility, such as E484K. Similar to N501Y, E484K increases binding affinity to the ACE-2 receptor [63].
The Delta variant quickly became the dominant strain of SARS-CoV-2 in India during the first half of 2021. As of 27 July, 2021, the Delta variant has been reported in 132 countries (Fig. 2) [1]. The spike protein of the Delta variant contains three key mutations: L452R, E484Q, and P681R.
These mutations can result in resistance to neutralizing monoclonal antibodies, favor antibody escape, and increase transmissibility by increasing the viral load, respectively [64]. It has been estimated recently that the Delta variant is 41% more transmissible than the Alpha variant. In a previous report, this value was overestimated (60% and 50%) [65]. In addition, the Delta variant carries a higher risk of hospitalization than the Alpha variant.
According to the COVID-19 Weekly Epidemiological Update, the reproduction number relative to non-VOCs was estimated to be 43–93% (95% CI 38–130%), 36% (95% CI 32–40%), and 11% (95% CI 7–16%) for the Alpha variant [66]. The Scientific Advisory Group for Emergencies (SAGE) estimated the R value to be 40–80% higher for the Delta variant than for the Alpha variant, using multiple modeling [65].
The B.1.427 and B.1.429 (West Coast) variants were first reported in the US state of California, with a similar 20% increased transmissibility compared to circulating non-B.1.427/B.1.429 lineages. The increased transmissibility may be due to a twofold higher viral load, detected in both infected patients and in in vitro experiments. There are no data about the transmissibility of the B.1.525 (UK and Nigeria) and B.1.1.207 (Nigeria) variants [30, 67, 68].
Pereira reported that SARS-CoV-2 variants combining spike mutations with a lack of ORF8 may be associated with a prolonged infection period, increased virus transmissibility, and less-severe disease [28]. There have been several studies suggesting that the ORF8 protein is involved in immune evasion. There have also been several studies showing that the E484K mutation may be linked to resistance to neutralization by antibodies [30].
The spike protein of B.1.617 contains L452R and E484Q mutations in the RBD region and a P681R mutation in the polybasic cleavage site [69, 70], which may accelerate the spread of the B.1.617 lineage worldwide. The mutation in the furin cleavage site (P681R) can enhance transmissibility due to an increased rate of membrane fusion and viral entry [43].
Based on epidemiological and modeling data, the Delta variant seems to have a transmission advantage over previously circulating variants [69, 71].
Immune escape
As the infection spreads and the number of immune people grows, the virus gains the ability to evade naturally induced immunity, having a chance to reinfect people [72]. Monitoring of escape mutations should be considered a public health priority. At first, sequencing can be used, but once a variant is recognized as being “of concern”, a PCR-based assay or other screening method should be immediately designed, validated, and made accessible globally for efficient real-time monitoring of the variant’s distribution [72].
Past serological studies have shown that a typical individual is infected once every 2-4 years with one of the four common cold coronaviruses – HCoV-NL63, HCoV-229E, HCoV-OC43, or HCoV-HKU1. This indicates that while reinfection occurs at a low rate, immunity against common cold coronaviruses is not highly durable [73, 74].
Deletions in the SARS-CoV-2 spike protein frequently occur in so-called recurrent deletion regions (RDRs), which affect epitopes for antibodies. Ninety percent of RDRs are in one of four discrete sites within the NTD, numbered 1 to 4 from the 5′ to the 3′ end of the S gene. Some SARS-CoV-2 variants are RDR variants, including mink cluster 5 Δ69–70, Alpha variant Δ69–70, Δ144/145, and Beta variant Δ242–244. These RDRs confer resistance to neutralizing antibodies [75].
Regarding the D614G variant, in contrast to previous studies with convalescent sera that showed slightly increased neutralization of the D614G variant versus wild-type following natural infection [76], Garcia-Beltran et al. demonstrated that, in individuals who were administered two doses of either BNT162b2 or mRNA-1273 vaccines, D614G pseudovirus neutralization was comparable to that of the WT form, with a 1.2-fold decrease for both two-dose vaccines [77].
This discrepancy might be due to the fact that most of the convalescent patients in previous studies were probably infected with the globally dominant D614G variant at that time, while the vaccines contain the wild-type spike sequence. A 1.4- to 2-fold mean reduction in neutralization titer for the B.1.429 variant was also observed in individuals who received two full doses of either a BNT162b2 or an mRNA-1273 vaccine [77].
A recent study revealed that the Alpha variant remains sensitive to neutralization, albeit at moderately reduced levels (~twofold), by serum samples from convalescent individuals and recipients of an mRNA-1273 vaccine (Moderna) and a protein nanoparticle NVXCoV2373 vaccine (Novavax) [78].
These results are in line with others indicating the comparable neutralization capacity of serum panels obtained from convalescent patients and vaccinated individuals against the Alpha variant [77, 79, 80]. Taken together, these recent data revealed that pre-existing immunity against the wild-type spike protein of SARS-CoV-2, either through natural infection or vaccination, provides adequate protection against the Alpha, B.1.429, and B.1.1.298 variants.
Hoffmann et al. showed that soluble ACE2 (sACE2), membrane fusion inhibitors, and TMPRSS2 inhibitors are effective against SARS-CoV-2 and its important variants of concern, particularly the Alpha, Beta, and Gamma variants [81]. However, in the case of antibody therapy, broad cross-neutralization was not always observed. While all of the antibodies evaluated, including casirivimab, imdevimab, and bamlanivimab, inhibited entry and blocked wild-type SARS-CoV-2 infection, only imdevimab showed the same level of inhibition of cell entry driven by the S proteins of the Beta and Gamma variants.
In fact, the Beta and Gamma variants showed partial and complete resistance to casirivimab and bamlanivimab, respectively. However, a cocktail of monoclonal antibodies called REGN-COV-2, consisting of casirivimab and imdevimab, efficiently blocked all of the variants tested [81]. On the other hand, sera from convalescent patients and individuals who received a BNT162b2 vaccine showed full protection against wild-type SARS-CoV-2 and slightly less protection against the Alpha variant. Thus, the BNT162b2 vaccine may provide less-reliable protection against infection by the Beta and Gamma variants [81].
Interestingly, Edara et al. reported that despite a few-fold reduction, most infected individuals still had binding and neutralizing antibodies against the Beta variant 8 months after infection. Furthermore, all Moderna mRNA-1273-vaccinated individuals produced neutralizing antibodies against the Beta variant [82].
In contrast, 36.7% and 42.9% of individuals who received two doses of BNT162b2 and mRNA-1273, respectively, had no detectable neutralization against the Beta variant. A lack of neutralization of the Beta variant was more likely when only one dose of the mRNA vaccine had been administered [77]. Therefore, a single dose of the current mRNA vaccines might be insufficient to elicit a cross-neutralizing antibody response in previously uninfected individuals.
In another study, in comparison to the Victoria isolate, a Wuhan-related isolate, neutralization titers against the Beta variant were 13.3-fold lower in early convalescent samples. However, when sera from patients infected with the Alpha variant were tested, there was a 3.1-fold difference in neutralization titer between the Victoria isolate and the Beta variant [83].
The neutralization titers of sera from individuals vaccinated with either the Pfizer-BioNTech BNT162b2 or the AstraZeneca/Oxford AZD1222 vaccine were also tested, and the results showed a 7.6- and 9-fold lower geometric mean titer, respectively, for the Beta variant than for the Victoria isolate, [83].
Two of two monoclonal antibody (MAbs) cocktails, one containing MAbs REGN10933 and REGN10987, and one containing MAbs AZD1061 and AZD8895, from Regeneron and AstraZeneca, respectively, were tested against the Beta variant, and it was found that the mutations in the Beta variant had no impact on neutralization by REGN10987, but they negatively impacted neutralization by REGN10933 (773-fold).
In contrast, no difference was observed with AZD1061 and AZD8895 [83]. The AZD7442 cocktail, which targets the RBD region, is a combination of two long-acting antibodies (LAABs) generated from B cells from convalescent patients: tixagevimab (AZD8895) and cilgavimab (AZD1061) [84]. Consistent with these data, several other studies have shown a reduction in cross-neutralization of the Beta variant by convalescent and vaccinated sera [77, 80, 85, 86].
A neutralization test using the B.1.427/B.1.429 variant revealed a 4.0- to 6.7-fold reduction in neutralizing titers in sera obtained from convalescent patients in comparison to wild-type pseudoviruses, while a 2.0-fold reduction was observed in mRNA vaccine recipients [16]. This is inconsistent with other studies that showed a reduction in the neutralization titer against the B.1.427/B.1.429 variant [87,88,89].
In the receptor-binding motif (RBM) of the spike protein, the B.1.427/B.1.429 (CAL.20C) and CAL.20A variants have an L452R substitution. According to one study, 36% of people who tested positive for SARS-CoV-2 after one or two doses of mRNA vaccine were positive for the L452R mutation, suggesting infection by the B.1.427 or B.1.429 variant [90]. Although vaccination resulted in better protection against the B.1.427 and B.1.429 variants than did natural infection, neither vaccination nor natural infection could confer complete protection against these variants. Presumably, due to the rapid changes occurring in the SARS-CoV-2 spike protein, this protection will be too weak to be protective in the future.
Regarding ACE2 mimics, the B.1.427 and B.1.429 variants showed moderate resistance to peptide-based inhibitors such as LCB1, and since the L452R mutation is located near the ACE2-LCB1 interface, it might affect the binding affinity of LCB1 [87, 91]. In addition, due to an S13I mutation in the signal peptide cleavage site that causes a significant structural rearrangement of the NTD antigenic supersite, a complete loss of B.1.427/B.1.429 neutralization was observed with a panel of mAbs targeting the NTD [87].
Another investigation showed that despite the B.1.617.1 variant being 6.8-fold more resistant to neutralization by sera from Moderna- and Pfizer-vaccinated subjects, the B.1.617.1 variant was neutralized by the majority of sera from convalescent patients and all sera from vaccinated individuals [92]. Despite in vitro data showing only a small decrease in neutralization against B.1.617 with the Covaxin vaccine, another study demonstrated vaccine breakthrough in healthcare workers who were fully vaccinated with the ChAdOx-1 vaccine [69, 93].
Regarding vaccine effectiveness (VE) of available vaccines against VOCs, NVX-CoV2373 has shown 86%, and 60% efficacy against Alpha and Beta, respectively [94]. The VE of AZD1222 was 70% against Alpha [95]. The VE after two doses of BNT162b2 mRNA vaccine was estimated to be 89% against Alpha [96], 86% against severe infection with Beta [97], 84% against Beta and Gamma [96], and 87% against Delta [96]. However, the VE after two doses of mRNA-1273 mRNA vaccine was 92% and 96.4% against Alpha [96] and Beta [98], respectively. Another study estimated the VE of full vaccination with the BNT162b2 mRNA vaccine to be around 84.4%, for Alpha and 64.6% for Delta in Norway [99]. Another study showed the VE of the Gam-COVID-Vac (Sputnik V) vaccine to be 78.6% in elderly people in Argentina for preventing confirmed infections and 87.6% and 84.8% for reduction in hospitalizations and deaths, respectively [100].
A series of studies demonstrated that the monoclonal antibodies LY15 CoV555, CB6, REGN10989, REGN10933, and REGN10987 provided a high potency of neutralization against the Alpha variant (IC50, 0.0001-0.0401 μg/mL) but LY-CoV555, CB6, REGN10989, and REGN10933 could not neutralize the Beta variant (IC50 >10 μg/mL) [85, 101, 102]. In a study measuring levels of IgA and IgG in vaccinated and convalescent saliva samples, convalescent individuals had significantly higher levels of IgA than vaccinated individuals (P = 0.03). Conversely, the IgG response in the saliva of vaccinated individuals was significantly higher than in saliva from convalescent individuals (P < 0.0001) [103].
Once selective pressure is exerted in the context of drug treatment, any drug resistance becomes more evident. Several lines of evidence strongly support the idea that cocktail therapy may be a powerful tool for controlling SARS-CoV-2 mutational escape, particularly when two antibodies bind to distinct and non-overlapping regions of the viral target, such as the RBD, making it unlikely that simultaneous mutations at two different genetic sites will allow viral escape. As a consequence, a therapeutic antibody cocktail may not only treat the disease but also protect against the development of resistance to antibody treatment [102].
L452R and E484Q are mutations in the receptor-binding domain of the spike, which not only binds ACE2 but also serves as a target for neutralizing antibodies. Pseudotyped viruses containing spike proteins of the B1.617.1 sublineage with three signature mutations (L452R, E484Q, and P681R) exhibited moderately decreased sensitivity to antibodies elicited by BNT162b2 mRNA vaccine, comparable to the loss of sensitivity conferred by L452R or E484Q alone [69]. In India, several people who were vaccinated with the Covishield vaccine were infected with the B.1.617 variant. Neutralizing antibodies elicited by natural infection or as a result of vaccination were found to be less effective against the B.1.617 variant [69, 104].
The N501Y and E484K mutations, which have been reported recently in VOCs, are located within the RBD. The N501Y mutation, which is present in nearly all of the major circulating VOCs (i.e., the Alpha, Beta, and Gamma variants), has no adverse effect on the neutralizing ability of sera from either naturally infected or mRNA-1273-vaccinated individuals [57, 82, 105, 106]. In contrast, the E484K substitution, which is present in the Beta and Gamma variants, is associated with resistance to conval
escent sera [86, 106,107,108]. Reinfection with SARS-CoV-2 has also been correlated with the E484K mutation [109,110,111], which can help the virus to evade polyclonal antibody responses [112]. Some research has also shown the K417N mutation to play a role in immune escape, especially in evading neutralization by a class of VH3-53/66 antibodies (Table 3) [86].
Table 3 The signature mutations of VOCs and their functional characteristics
From: The ins and outs of SARS-CoV-2 variants of concern (VOCs)
| Signature mutation | Region | Function | Ref. |
|---|---|---|---|
| HV69/70del | NTD | May affect recognition by antibodies, located in a “recurrent deletion region”, diagnostic real-time PCR detection failure | [24, 78, 156] |
| k417N | RBD | May escape antibody binding and decrease ACE2 binding | [157, 158] |
| L452R | RBD | May augment ACE2 binding, increase antibody neutralization, and permit cellular immunity escape | [159, 160] |
| S477N | RBD | May increase ACE2 binding and reduce neutralization by antibodies and convalescent sera | [161, 162] |
| E484K | RBD | May reduce convalescent serum neutralization (immune escape) and increase ACE2 binding. It has been associated with reinfection. | [64, 163,164,165] |
| N501Y | RBD | May increase ACE2 binding and antibody recognition | [64] |
| D614G | Spike | May increase infectivity | [163] |
| P681H | Spike | May reduce antibody recognition, be important for immune recognition, and increase the rate of S1-S2 cleavage | [159, 159, 159] |
N439K is a mutation that can help the virus to evade antibody-mediated immunity while maintaining fitness in comparison to WT. The N439K RBD retains high-affinity interactions with the hACE2 receptor, and viruses containing the N439K mutation replicate efficiently in cultured cells. Furthermore, no difference in disease severity has been associated with this mutation. Evasion of antibody-mediated immunity was assessed by testing the recognition of N439K RBD by mAbs and polyclonal sera from 442 convalescent patients [113].
The second arm of the immune response is cellular immunity. T cell epitope substitutions may impair cytotoxic T lymphocyte or T helper recognition, resulting in delayed elimination of infected cells or suboptimal B cell and antibody response assistance. T cell epitopes, unlike neutralizing antibody epitopes, are found in the entirety of the spike protein. It is not yet clear how mutations affect T cell responses or lead to T cell escape variants. However, the polyclonal response of T cells targeting several regions of the S protein of SARS-Co-V-2 might limit the influence of mutations on cellular immunity [114].
Furthermore, for circulating variants such as Beta and Gamma, it is reasonable to predict that T-cell-mediated immunity elicited by infection would be protective. Motozono and colleagues found that the L452R and Y453F mutations in the B.1.427/B.1.429 and B.1.298 variants contribute to escape from HLA-A24-medicated cellular immunity [115]. This implies a role of CTL responses in the control and evolution of SARS-CoV-2 and vice versa. Another study showed no difference in CD4+ T cell activation in response to the Alpha and Beta variants [116]. Furthermore, the presence of SARS-CoV-2-specific memory T cells has been detected 10 months after infection [115, 117].
There are also different strategies that SARS-CoV-2 uses to escape the innate immune response [118]. It has been demonstrated that SARS-CoV-2 is sensed by innate immune sensors such as TLR-3 and TLR-7 [119, 120] and that the Nsp8, ORF9b, N, and M proteins of SARS-CoV-2 are involved in escape from innate immune sensors. Nsp8 suppresses the MDA5-dependent IFN signaling pathway by direct binding to MDA5 CARD and blocking K63-linked polyubiquitination [121].
ORF9b antagonizes the type I IFN pathway via association with translocator of outer membrane 70 (TOM70), a critical mitochondrial import receptor regulating IFN responses [122]. A recent study showed that SARS-CoV-2 N suppresses the phosphorylation and nuclear translocation of STAT1 and STAT2, thus interfering with the IFN signaling pathway [123]. Moreover, another study has shown that the SARS-CoV-2 M protein suppresses type I and III IFN production. SARS-CoV-2 M binds directly to essential molecules of the cytosolic viral RNA sensing pathway, such as RIG-I, MDA5, MAVS, and TBK1, and prevents their interaction [124].
Risk of hospitalization and mortality
VOCs can spread quickly in communities, leading to an increased number of COVID-19 patients, a higher hospitalization rate, increased pressure on healthcare systems, and higher mortality rates, mainly in older individuals with comorbidities [49]. A matched cohort study was performed in the UK by Challen et al. to evaluate changes in mortality rates related to the Alpha variant. The mortality hazard ratio was estimated to be 1.64 (95% CI 1.32–2.04) in patients who tested positive for novel variants, with 2.5–4.1 deaths per 1000 confirmed cases [66]. Davis et al. estimated a 61% higher mortality rate for B.1.1.7 than for the previous variants [125]. Furthermore, B.1.1.7 was associated with a ??relative risk?? (RR) of hospital admission of 1.42 (95% CI 1.25–1.60; p < 0·0001) in Denmark. Moreover, the in-hospital mortality risk was estimated to be 20% higher for the Beta variant (adjusted OR: 1.2, 95% CI 1.2–1.3) [50].
A meta-analysis data showed a strong correlation between age and infection fatality rate (IFR) in COVID-19 cases. The age-specific IFR was estimated to be 0.002%, 0.01%, 0.4%, 1.4%, 4.6%, and 15% at the age of 10, 25, 55, 65, 75, and 85, respectively [126]. There are also differences in mortality among different racial and ethnic groups. For instance, an analysis of the COVID-19 mortality cumulative incidence (CmI) demonstrated that Blacks/African Americans have 34% of the total mortality in the United States despite representing only 13% of the population. The COVID-19 CFR was also higher among Blacks/African Americans relative to Whites [127]. No peer-reviewed articles are available for the CFR of the Delta variant.
A higher hospitalization risk was observed in American patients infected with Gamma (HR 3.17, 95% CI 2.15–4.67), Beta (HR: 2.97, 95% CI 1.65–5.35), Delta (HR: 2.30, 95% CI 1.69–3.15), and Alpha (HR 1.59, 95% CI 1.26–1.99) than in those infected with a non-VOC lineage [128]. The hospitalization risk after infection with the Delta variant has also been reported for a cohort in Scotland (HR 1.85, 95% CI 1.39–2.47) [129] and England (HR 2.61, 95% CI 1.56–4.36) [130]. In a study conducted in Singapore, the Delta variant was associated with indicators of higher disease severity, such as oxygen requirement, ICU admission, or death (adjusted odds ratio (AOR) 4.90, [95% CI 1.43–30.78]) [131].
Diagnostics
Specific detection of variants is done by various methods [132]. Laboratories may choose one or more methods based on the facilities and technology available, their contacts with partner laboratories, and the number of trained specialists, as well as their epidemiological goals, such as disease control and monitoring [30, 133, 134]. These methods include complete sequencing of the viral genome, partial gene sequencing, and real-time-PCR-based screening techniques [135].
Sequencing is the gold standard and is a reliable and accurate method for identifying variants. In this case, the whole viral genome is sequenced by a tailed-amplicon approach or shotgun sequencing, and the results are compared with other data in SARS-CoV-2 databases [136]. This method can be used as a reference method without prior knowledge of specific mutations in the viral genome. Partial next-generation sequencing or Sanger sequencing is an alternative way to identify VOCs [137] by limiting sequencing to the S gene.
Although this technique shares the disadvantages of whole-genome sequencing in terms of high cost and the need for sophisticated bioinformatics analysis and advanced equipment, it can be done more quickly. Several multiplex and allele-specific real-time-PCR-based techniques have been proposed for screening variants [135]. In the spike Δ69-70 screening test, S gene target failure (SGTF) is seen with some VOCs (such as the Alpha variant) due to the deletion [136].
The TaqPath commercial multiplex real-time PCR kit, which simultaneously identifies three genes – N, ORF1ab, and S – is affected by SGFT but can still be used to screen for variant B1.1.7 [138]. It should be noted that SGFT is observed not only with the B1.1.7 variant but with other variants as well.
As an alternative to sequencing, mutation screening tests have been developed. For example, Vogels et al. designed a screening test for the Alpha, Beta, and Gamma variants in which two distinct regions including Δ3675-3677 of ORF1ab and Δ69-70 of S are targeted in a real-time-PCR-based assay [139]. The ORF1ab gene deletion is seen in all three of the above-mentioned VOCs, whereas the 69-70 gene deletion is seen only in the UK variant.
Multiplex S gene Δ69-70 and N501Y real-time PCR is also used as a practical test to screen for VOCs. The difference between this method and the test described above is that N501Y has been selected as a common target in the Alpha, Beta, and Gamma VOCs [140]. The N501Y screening test detects N501Y in three variants of concern by allele-specific PCR [141].
In the N501Y and A570D screening test, A570D is present only in the Alpha variant, whereas N501Y is present in the Alpha, Beta, and Gamma variants [142]. An E484K screening test is designed to detect the E484K amino acid change in the P1 and B1.135 variants and in some B1.1.7 variants that also have this mutation [143]. In addition, some commercial screening kits detect variants by analyzing all of the significant mutations using real-time PCR followed by melting curve analysis [144].
Omicron variant of concern (VOC)
This weekly epidemiological update provides an overview of the extent to which SARS-CoV-2 Omicron VOC is prevalent in EU/EEA countries, based on the best currently available evidence (as of 20 January 2022). The data are collected from The European Surveillance System (TESSy) or the GISAID EpiCoV database. Please refer to the ECDC Variants of interest and concern in the EU/EEA dashboard for more details. In cases of missing data, EU/EEA countries’ official national or regional websites are used, which are linked to the country name below. As a result, the data presentation and completeness might be different depending on the data sources and availability.
As of 20 January 2022, the Omicron variant has been identified in all EU/EEA countries. From 20 December 2021 to 9 January 2022, 23 EU/EEA countries with adequate sequencing volume reported an estimated prevalence of Omicron of 69.4% (range from 5.7% to 99.9%), over 20% higher than in the previous week.
Countries where Omicron has become the dominant variant (accounting for more than 50% of sequenced viruses) include Austria (95.4%, 2022-02), Belgium (99.7%, 2022-02), Cyprus (93.9%, 2022-01)*, Czechia (66.7%, 2022-02), Denmark (98.8%, 2022-02), Finland (99.9%, 2022-02), France (90.8%, 2022-02), Germany (62.5%, 2022-01), Greece (85.6%, 2022-01), Hungary (64.7%, 2022-02), Iceland (90%, 6 January 2022), Ireland (89.2%, 2021-52)*, Italy (81%, 2022-01), Liechtenstein (88.5%, 2022-01)**, Lithuania (40.5%, 2021-52)*, Luxembourg (89.6%, 2022-01), Malta (99.3%, 2022-01)*, the Netherlands (95.3%, range between labs 93.3% – 98.6%, 10 January 2022), Norway (93.8%, 2022-02), Portugal (86.3%, 2022-01), Slovenia (67% of cases sequenced on 5 January 2022), Spain (87.4%, 2022-01), and Sweden (91.8%, 2022-01).
Countries where the Omicron variant is present but not dominant include Bulgaria (43.5%, 2022-02), Croatia (no national proportion available), Estonia (45.8%, 2022-02), Latvia (5.8%, 2022-02), Poland (26.2%, 2022-02), Romania (37.8%, 2022-02), and Slovakia (29.9%, 2022-02).
Large (20% or higher) increases in the number of cases since last week have been reported in the following countries: Bulgaria, Cyprus, Czechia, Estonia, Germany, Hungary, Italy, Luxembourg, Malta, Spain, and Sweden. For a general overview of the epidemiological COVID-19 situation in the EU/EEA, please see the ECDC weekly Country Overview ReportECDC Country Overview Report.
Based on case-based data, 155 150 Omicron cases were reported to TESSy between weeks 2021-46 and 2022-02 from: Austria (number of cases 84 537; proportion of all cases 54%), Cyprus (343; 0%), Estonia (27; 0%), Finland (8 149; 5%), Ireland (1 404; 1%), Italy (3 231; 2%), Liechtenstein (35; 0%), Luxembourg (974; 1%), Malta (296; 0%), Norway (50 534; 33%), Poland (37; 0%), Portugal (570; 0%), Romania (92; 0%), Slovakia (112; 0%), and Sweden (4 809; 3%).
- The median (interquartile range) age of these Omicron cases was 30 (20–33) years, 7% were aged 60 years and above, and 50% were male.
- Among 111 946 (72%) Omicron cases with complete data on symptom status, 84 662 (76%) cases were reported as symptomatic and 27 284 (24%) as asymptomatic.
- Complete data on vaccination status were available for 2 369 (2%) Omicron cases, among which 211 (9%) were reported as one dose, 1 646 (69%) were reported as two doses, 255 (11%) were reported as three doses, 239 (10%) were reported as unvaccinated, and 18 (1%) were reported as vaccinated (unknown doses).
- Among 124 849 (80%) Omicron cases with complete data on importation status, imported or travel-related cases accounted for 9 159 (7%) cases and 115 690 (93%) cases were locally acquired.
- Among Omicron cases with known outcomes, 884 (1.14%) were hospitalised, 120 (0.16%) required ICU admission/respiratory support, and 48 (0.06%) died. The status of COVID-19 outcomes was known for 77 233 (50%) cases for hospitalisation, 76 744 (49%) for ICU admission/respiratory support, and 81 912 (53%) for death (some incomplete reporting of these variables is expected for more recent cases).
*Insufficient precision at less than 5% prevalence (more information available here).
** Seven-day average. Please note that the data for Liechtenstein are reported together with Swiss data.
Disease severity related to Omicron
Evidence from a variety of settings suggests that infections with the Omicron VOC have a less severe clinical presentation than those due to the Delta VOC. It is important to highlight that lower age, prior immunity from natural infection, vaccination including booster dose, and improved treatment options will contribute to less severe outcomes from subsequent infection. Therefore, the true risk of severe infection for Omicron may be underestimated by the large numbers of vaccinated or previously infected people, which was not the case in the beginning of preceding waves. Most studies do not account for waning immunity, or for the likely large amount of under-ascertained reinfections. This could lead to an underestimation of severity.
Among Omicron cases with known outcomes reported to TESSy as of 19 January 2022, 884 (1.14%) were hospitalised, 120 (0.16%) required ICU admission/respiratory support, and 48 (0.06%) died. The pattern of higher rates of hospitalisation, ICU admission, and death with increased age is apparent for Omicron cases, as it has been for Delta and previous variants. Similarly, low hospital admission rates (0.3%) and case fatality (<0.1%) for Omicron cases have been observed in Canada, and in Texas, California, and Denmark a shorter median length of hospital stay and/or significantly reduced need for respiratory support were reported for Omicron. This observed severity in TESSy data and in the studies cited is at least partially likely due to the protective effect of vaccination, time since vaccination, co-morbidities, and/or previous infection in some of the individuals and does not necessarily reflect the inherent severity of Omicron.
Across studies from various settings, the risk of hospitalisation was found to be lower for Omicron than for Delta or other previous variants. While studies used slightly different data, analysis approaches, and adjustments for confounding factors, most studies found risk reduction in the range of 50-60%.
Preliminary analysis of case-based data submitted by 15 EU/EEA countries to TESSy between week 46-2021 and week 02-2022 was performed to compare the overall adjusted odds ratio (aOR) of hospital admission for infection with Omicron compared to infection with Delta among symptomatic cases. Logistic regression models adjusted for age group, sex, preconditions, reporting country, reporting week and vaccination status showed that Omicron infection was less likely to be reported with admission to hospital compared to infection with Delta (aOR 0.41; 95% CI: 0.37-0.46). More in-depth analysis will be performed over the course of the upcoming weeks to better assess the impact of Omicron on severe outcomes reported to TESSy.
A Danish observational cohort study of 188 980 SARS-CoV-2 positive individuals during November-December 2021 compared the risk ratio of admission for Omicron compared to Delta infection and stratified by vaccination status and found that Omicron was associated with an adjusted RR of hospitalisation of 0.64 (95% CI: 0.56-0.75) compared to Delta infection. RR was 0.57 (95% CI: 0.44-0.75) among cases with none or one vaccination, 0.71 (95% CI: 0.60-0.86) among two-dose vaccinated, and 0.50 (95% CI: 0.32-0.76) among three-dose vaccinated. Similarly, Omicron had lower risk than Delta cases with regard to vaccination status, age, comorbidity, and reinfection. The lower hospitalisation risk for Omicron cases among both vaccinated and unvaccinated individuals suggests a reduced severity of Omicron, but Omicron patients in this study were younger and had fewer comorbidities than those with Delta.
In the United Kingdom, after adjusting for sex, age, travel, vaccination status and, where ascertained, previous infection, presentation to emergency care or hospital admission with Omicron was approximately half of that for Delta (Hazard Ratio 0.53, 95% CI: 0.50 to 0.57). They also found a 65% lower hospitalisation risk for Omicron cases who had received two doses of a vaccine and 81% reduction with three doses, compared to unvaccinated Omicron cases.
In Scotland, symptomatic individuals who were S-gene negative (a proxy for Omicron infection) had a two-thirds reduced hospitalisation risk compared to S-gene positive (Delta) cases, while the rate of possible reinfection for Omicron was 10 times that of Delta. People who had received three vaccine doses had a 57% (95% CI 54-60) lower risk of experiencing symptoms following Omicron infection compared to Delta.
In Southern California, the United States, the risk of hospital, ICU admission, and mortality were 0.48 (0.36-0.64), 0.26 (0.10-0.73) and 0.09 (0.01-0.75) higher, respectively, among cases with Omicron infection compared to cases with Delta infection.
It usually takes several weeks for the accumulation of clinical outcomes to make conclusions on the impact of a specific variant on hospital admissions, intensive care needs, and death rates. It is also essential to account for the relatively young age of most people who have been infected with Omicron to date, and thus far there are little data on the severity among older age groups and people with underlying risk factors. As a result, the clinical profile of Omicron may change as other groups are infected and followed over time.
Significantly, the combination of a higher growth rate and immune evasion indicate that any potential advantage Omicron may have in terms of decreased severity might be countered by increased community infection rates that lead to a substantial additional burden for hospitals, while primary care may be overburdened even more than in previous waves. As more evidence builds up, a better assessment of clinical outcomes and long-term consequences, such as post-COVID-19 condition, will be feasible.
Vaccine effectiveness against infection and/or severe disease due to Omicron
A recent (pre-print) study from Denmark estimated the transmission dynamics following the spread of Omicron within Danish households during December 2021. The study found the Secondary Attack Rate (SAR) to be 31% and 21% in households with the Omicron and Delta variants, respectively. Unvaccinated potential secondary cases experienced similar attack rates in households with the Omicron and Delta variants (29% and 28%, respectively), while fully vaccinated individuals experienced secondary attack rates of 32% in household with Omicron and 19% in households with Delta. For people who had received a booster dose, Omicron was associated with a SAR of 25%, while the corresponding estimate for Delta was 11%. There was an increased transmission for unvaccinated individuals, and a reduced transmission for booster-vaccinated individuals, compared to fully vaccinated individuals. Comparing households infected with Omicron to Delta, the study found a 1.17 (95% CI: 0.99-1.38) times higher SAR for the unvaccinated, 2.61 times (95% CI: 2.34-2.90) higher for the fully vaccinated, and 3.66 (95% CI: 2.65-5.05) times higher for booster-vaccinated individuals, indicating immune evasiveness of the Omicron variant.
The United Kingdom’s Health Security Agency (UKHSA) provides regularly updated estimates of vaccine effectiveness against symptomatic infection with Omicron, and has reported estimates of 63% (95% CI: 59%, 67%), 68% (95% CI: 55%, 78%), and 25% (95% CI: 1%, 43%), in the initial period (two to four weeks) after primary vaccination with Comirnaty, Spikevax and Vaxzevria, respectively. From 25 weeks after the second dose, the vaccine effectiveness decreased to low or non-significant levels but was restored after a booster dose of Comirnaty to 69% (95% CI: 67%, 70%) for those who had received a primary course of Comirnaty, and to 64% (95% CI: 63%, 66%) for those who had received Vaxzevria as primary vaccination. UKHSA has also conducted a separate analysis of individuals aged 65 years and older and found that, in all periods after vaccination, effectiveness was lower for Omicron compared to Delta. From 20 weeks after the second dose of either Vaxzevria or Comirnaty, minimal or no effect against mild disease was seen for the Omicron variant. Although a booster dose of either Comirnaty or Spikevax temporarily increased the protection, this also waned with time, to around 30% vaccine effectiveness against mild disease at 10+ weeks following a booster dose.
A preprint of a study from Canada assessing the vaccine effectiveness of mRNA vaccines found that two doses of an mRNA vaccine were unlikely to protect against Omicron infection (VE 6%, 95% CI: -25, 30%). A third dose provided some protection against Omicron infection (VE 37% (95% CI: 19%, 50%) in the immediate term but substantially less than against Delta (VE 93% (95% CI: 92%, 94%), ≥7 days after third dose. The study highlights that the results may be confounded by behaviours that were unable to be accounted for and that the duration of the protection and effectiveness against severe disease are uncertain.
Another preprint study from the United States, contracted by the vaccine manufacturer Moderna, reports a vaccine effectiveness of 30.4% (95% CI: 5.0%, 49.0%) against Omicron infection after two doses of the Spikevax vaccine. A third (booster) dose increased the effectiveness to 62.5% (95% CI: 56.2%, 67.9%). The study also reports considerably lower vaccine effectiveness against Omicron infection among immunocompromised individuals compared to the general population.
Earlier studies from the UK and Denmark showed significantly reduced and declining vaccine effectiveness against symptomatic disease with Omicron compared to infection with Delta. These studies also reported that a booster dose increased the vaccine effectiveness, although not as efficiently as against infection with Delta. The additional protection from booster doses against infection with Omicron was also confirmed in a recent study from Scotland.
Estimates of vaccine effectiveness against hospitalisation from the UK and South Africa have indicated that the protection against severe disease from Omicron infection is higher (around 70% after primary vaccination) than the protection against mild infections, and that the protection increases after a third vaccine dose.
The estimates of vaccine effectiveness against Omicron should still be considered as preliminary evidence, and more data are awaited from larger populations that allow for stratification of data by age, vaccine type, number of doses received, and time since vaccination. Studies and collection of real-life data are ongoing to further assess the level of protection from the vaccines against transmission, infection, and severe disease.
Overall level of risk and options for response
Based on the current situation and the available evidence, ECDC’s Rapid Risk Assessment (18th update) on the impact of Omicron remains valid: the overall level of risk to public health associated with the further emergence and spread of the SARS-CoV-2 Omicron VOC in the EU/EEA is assessed as VERY HIGH.
The very high growth advantage of Omicron has resulted in very high case notification rates. Such high levels of SARS-CoV-2 transmission leads to high levels of absence from work, including among healthcare and other essential workers, and likely overwhelm the testing and contact tracing capacities in many EU Member States. The sheer volume of COVID-19 cases places considerable strain on healthcare systems and society. ECDC has provided options for adapting quarantine and isolation, particularly when countries face high or extreme pressure on healthcare systems and other functions in society, including essential services.
Please see the ECDC’s Rapid Risk Assessment for epidemiological forecasts and options for response (non-pharmaceutical interventions, health system strengthening, vaccination, testing and contact tracing, and risk communication). ECDC will publish updates on the epidemiological situation, severity, spread, and vaccine effectiveness in short intervals.
Awaiting the availability of further evidence, urgent and strong action is needed to reduce transmission, keep the burden on healthcare systems manageable, and protect the most vulnerable.
Member States should urgently assess their acceptable levels of residual risks, current healthcare system capacities, and available risk management options (e.g. contingency and business continuity measures, surveillance and testing strategy, quarantine and isolation policy, etc.).
The continuation and strengthening of non-pharmaceutical interventions is necessary to reduce ongoing Delta and Omicron transmission and keep the COVID-19-related disease burden manageable. These measures include avoiding large public or private gatherings, extended use of face masks, reduced contacts between groups of individuals in social or work settings, teleworking, and reduced inter-household mixing.
Vaccination remains a key component of the multi-layered approach needed to address the ongoing circulation and reduce the impact of the Delta and Omicron variants. Efforts should continue to increase full vaccination uptake in individuals who are currently unvaccinated or partially vaccinated and accelerate the roll-out of booster doses.
Member States are strongly encouraged to conduct and share findings on outbreak investigations and epidemiological studies to inform future risk assessments.
reference link : https://www.ecdc.europa.eu/en/news-events/weekly-epidemiological-update-omicron-variant-concern-voc-week-2-data-20-january-2022
REFERENCE LINK : https://link.springer.com/article/10.1007%2Fs00705-022-05365-2



{kind=link}
[…] SARS-COV-2 : Current situation on Variants of Concern (VOCs) […]