











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Further they also found that these prion-like domains vary in the various SARS-CoV-2 variants, contributing to their difference in binding affinity.
Prions are proteins that possess a unique conformational conversion, with the ability to rapidly shift between multiple conformations due to residue hydrophobicity and net sequence charge, and viral prion-like proteins are known as potential regulators of viral infections.
However, the prion-like domains (PrD) in the SARS-CoV-2 proteome have not been analyzed.
The study findings were published in the peer reviewed journal: Microorganisms.
https://www.mdpi.com/2076-2607/10/2/280/htm
SARS-CoV-2 is a new member of the Betacoronavirus (β-CoV) genus of large, enveloped, single-stranded RNA viruses [2]. This genus not only includes viruses that cause deadly human infections such as severe acute respiratory syndrome (SARS) and Middle East respiratory syndrome (MERS) but also encompasses viruses that cause non-life-threatening common colds, including human coronavirus OC43 (HCoV-OC43) and human coronavirus HKU1 (HCoV-HKU1) [3].
Although these viruses predominantly infect lung epithelial cells, the clinical severity and pathogenesis of the infections they cause vary between different coronaviruses [4]. While severe pneumonia and pulmonary fibrosis are fundamental to the pathogenesis of COVID-19, SARS and MERS, these symptoms are not typical of infections caused by HCoV-OC43 and HCoV-HKU1 [5,6].
The key determinant of the host specificity of β-CoVs is the surface-located S protein, which plays critical roles in infection by mediating viral attachment to host cell-surface receptors and facilitating viral entry [8]. The S protein consists of two large regions: the N-terminal S1 and C-terminal S2 [9]. S1 is responsible for recognizing host-cell receptors, including the receptor-binding domain (RBD), and has higher sequence variability than S2 (S1 shares around 70% identity with that of other human β-CoVs).
The membrane-embedded S2 region responsible for fusion is more highly conserved than that of S1 [8,9]. In SARS-CoV-2, the RBD in S1 allows the virus to bind directly to the peptidase domain of the host angiotensin-converting enzyme 2 (ACE2) complex, mediating virus entry into sensitive cells [10].
Mutations in the genome sequence of SARS-CoV-2 are responsible for the emergence of new SARS-CoV-2 variants, many of which are characterized by higher transmission rates [15,16,17].
Recently, we conducted an analysis and identified for the first time viral prion-like domains (PrDs), which we suggest are novel regulators of virion assembly with a role to play in virus-host cell interactions [18,19]. These studies were in alignment with previous studies, showing that in addition to the pathological role prions play in humans—being implicated in Alzheimer’s and Parkinson’s diseases, diabetes, and many other human pathologies—protein misfolding also plays important physiological roles in eukaryotes and prokaryotes [20,21,22,23].
Though the detailed molecular mechanisms underlying prion formation remain elusive, asparagine (Q)- and glutamine (N)-rich regions characterized by altered hydrophobicity and net sequence charge are known to drive prion formation. This is the basis for a number of algorithms for identifying candidate prionogenic domains [24,25].
One such algorithm is prion-like amino acid composition (PLAAC) analysis, which allows for the evaluation of prion-like domains based on the hidden Markov model (HMM) that also incorporates other prion-like domain-predictive algorithms PAPA, DIANA, and FoldIndex [26,27,28].
Although structures for the variants of SARS-CoV-2 were extensively researched using CryoEM, modeling, molecular dynamics and other methods to study the impact on the binding affinity for each amino acid in contact with ACE2, the prionogenic properties of SARS-CoV-2 have not yet been studied [29,30,31,32].
In this study, we performed the first detailed evaluation of PrDs in the spike protein of SARS-CoV-2 and compared them to PrDs from other human-pathogenic β-CoVs. We also analyzed PrDs in the spike protein of the variants of concern (VOC) B.1.617.2 (Delta) and B.1.1.529 (Omicron), variants of interest (VOI) and variants being monitored (VBM), such as B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma), B.1.427 (Epsilon), B.1.617.1 (Kappa) and P.2 (Zeta), some of which are known for their ability to escape antibody neutralization [33,34,35,36].
Further analyses of these PrD-containing proteins in SARS-CoV-2 may improve our understanding of the COVID-19 infection and provide new insights into its pathophysiology and novel targets for developing therapies, including antiprion compounds with binding properties to prion proteins.

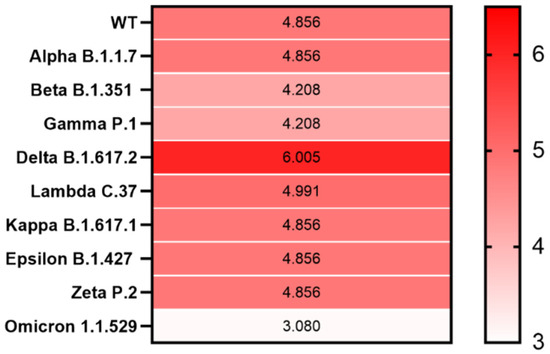
We analyzed the particularities of the PrDs in the SARS-CoV-2 variants, some of which are known to have a substantially increased binding affinity due to mutations in the S protein, thus suggesting that they may have greater prion-forming potential. To this end, we analyzed the correlation between the LLR score of the PrD within the RBD of the S proteins of the VOC, VOI and VBM.Compared with that of SARS-CoV-2 WT, we observed an elevated LLR score for the S protein from only the Delta (B.1.617.2) variant, with the LLR of 6.025, while from the emerging Omicron (B.1.1.529), the LLR was only 3.080 (Figure 3 and Supplementary Figure S4).



{kind=link}