









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Vervet monkeys with a strong preference for ethanol that were given an FGF21 analogue consumed 50% less alcohol.
The study also studied the brain circuits involved in mice and found that the protein, known to also reduce sugar intake, acts on different circuits to reduce alcohol and sugar consumption.
“When considering how and why these modality specific mechanisms evolved, it is interesting to note that mammals were primarily exposed to alcohol from fermenting fruits, which possess high levels of simple sugars,” says senior study author Matthew Potthoff of the University of Iowa Carver College of Medicine. “Despite this, neural circuits regulating FGF21-mediated suppression of sugar and alcohol intake apparently developed independently and not in response to a shared selective pressure.”
Recently, genome-wide association studies have shown that FGF21 genetic variants are linked to increased alcohol consumption in humans. In rodents, pharmacologic administration of this protein, which is produced in the liver, reduces alcohol consumption through actions in the brain.
But until now, the neural circuits through which FGF21 inhibits alcohol consumption were unknown, as were its effects on alcohol consumption in higher organisms.
In the new study, Potthoff and co-first author Kyle Flippo of the University of Iowa and international collaborators, including co-first authors Drs. Matthew Gillum and Samuel Trammell of the University of Copenhagen, showed that administration of an FGF21 analogue reduces alcohol intake by 50% in vervet monkeys with a strong innate preference for ethanol. FGF21 and the FGF21 analogue decrease alcohol intake even when administered after prolonged ethanol exposure in mice and primates.
FGF21 alters neural transmission in the nucleus accumbens, a brain region that plays a complex role in reward and addiction, and suppresses alcohol consumption through a sub-population of neurons in the basolateral amygdala.
Specifically, FGF21 signaling in neurons that project from the basolateral amygdala to the nucleus accumbens suppresses alcohol consumption by changing the activity of a specific subpopulation of these neurons.
Previous studies have shown that this pathway is involved in reward-seeking behavior. According to the authors, more research is needed to investigate the specific effects of FGF21 on the activity of these neurons during alcohol consumption in animal models.
“Our results provide a mechanism for a liver-to-brain endocrine feedback loop that presumably functions to protect the liver from damage,” Flippo says.
The Relationship Between FGF21, Metabolism, and Aging
FGF21 and Its Family
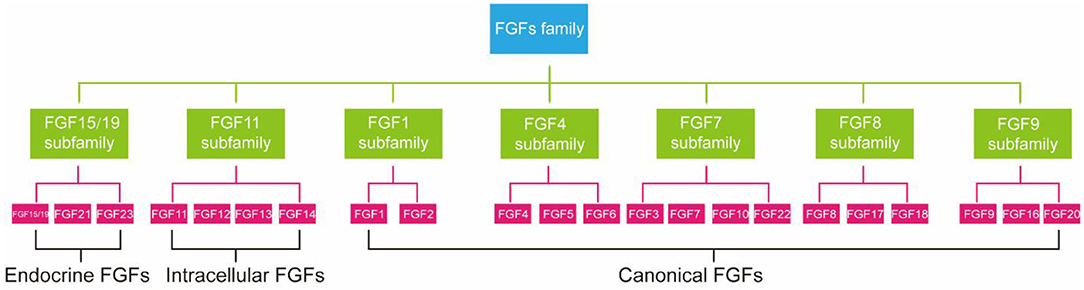
FGFs family consists of 23 members but only 18 FGFR ligands. Four family members (FGF11, FGF12, FGF13, and FGF14) cannot bind to FGFR are more correctly referred to as FGF homologous factors (38). FGFs are effective regulators of cellular aging. Not only that, mutations in FGFs have been linked to many metabolic diseases, including atherosclerosis, NAFLD, and diabetes (39). According to phylogenetic analysis, FGFs family members can be further divided into seven subfamilies (Figure 1) (40).
FGF21 is a member of an endocrine FGF subfamily, which includes FGF15/19 and FGF23. FGF21 can circulate and diffuse freely in tissues as an endocrine factor because of the lack of heparin-binding domain (41). FGF21 is expressed in many tissues, including liver (42), adipocytes (43), brown adipose tissue (BAT), pancreas (44), gastrointestinal tract, brain, skeletal muscle, and heart, and it directly regulates the metabolism and aging of peripheral tissues (12). Next, we will introduce the relationship between FGF21 metabolism and aging.
FGF21, the Hub Linking Metabolism and Aging?
In 2005, FGF21 was first used as a novel metabolic regulator (45). As a coordinator of energy metabolism in multiple organs (especially liver and fat), FGF21 can regulate adipogenesis, glucose uptake, and cellular insulin sensitivity (12, 46–48). The level of FGF21 is elevated to promote the oxidation of free fatty acids (FFAs) and inhibit lipogenesis in the liver to supply energy when glucose levels are low or caloric restricted (49). In an animal experiment, Inagaki et al. found that FGF21 expression increased 28-fold in the livers of mice after 12 h of fasting.
Increased FGF21 expression will stimulate ketogenesis in the liver and promote lipolysis in white adipose tissue to provide energy for activities (50). In addition to mice, in the fasting state, the level of FGF21 in human serum will also increase rapidly within a few hours to promote lipolysis (51).
Proper administration of FGF21 also effectively improved insulin sensitivity and hepatic glucose uptake in obese mice (52). Studies have found that long-term administration of FGF21 to genetically obese mice will ameliorate fasting hyperglycemia via increased glucose uptake and improved hepatic insulin sensitivity (53). Lack of FGF21 in mice evokes insulin resistance and promotes gluconeogenesis and liver glucose production (54). In addition to liver and adipose tissue, the expression of FGF21 in skeletal muscle also has important metabolic functions. FGF21 could improve muscular dystrophy and atrophy through metabolic pathways (55, 56).
In addition to acting as a metabolic regulator, FGF21 can also improve aging. There are clear indications that the effect of FGF21 in preventing aging may be related to the thymus; overexpression of FGF21 can prevent aging-related changes, such as retarding thymus degeneration to prevent thymus weakness, improving immune system, and hopefully extending human life expectancy in the future (57).
With aging, tissue autophagy is reduced, disrupting tissue ability to maintain protein homeostasis, thus accelerating the aging process (57). FGF21 can stimulate adiponectin secretion in fatty tissues, thus improving autophagy in target tissues to play an anti-aging role (58). Compared with normal mice, animal experiments show that fasting-induced FGF21 overexpression in mice and slowed aging (59). Transgenic overexpression of FGF21 significantly extended the life span of mice without reducing food intake or affecting NAD+ metabolism (60).
In summary, FGF21 mainly regulates aging by metabolism. It is crucial to clarify the mechanism or pathways of FGF21 in regulating metabolism and aging.
The Mechanism by Which FGF21 Regulates Aging Through Metabolism
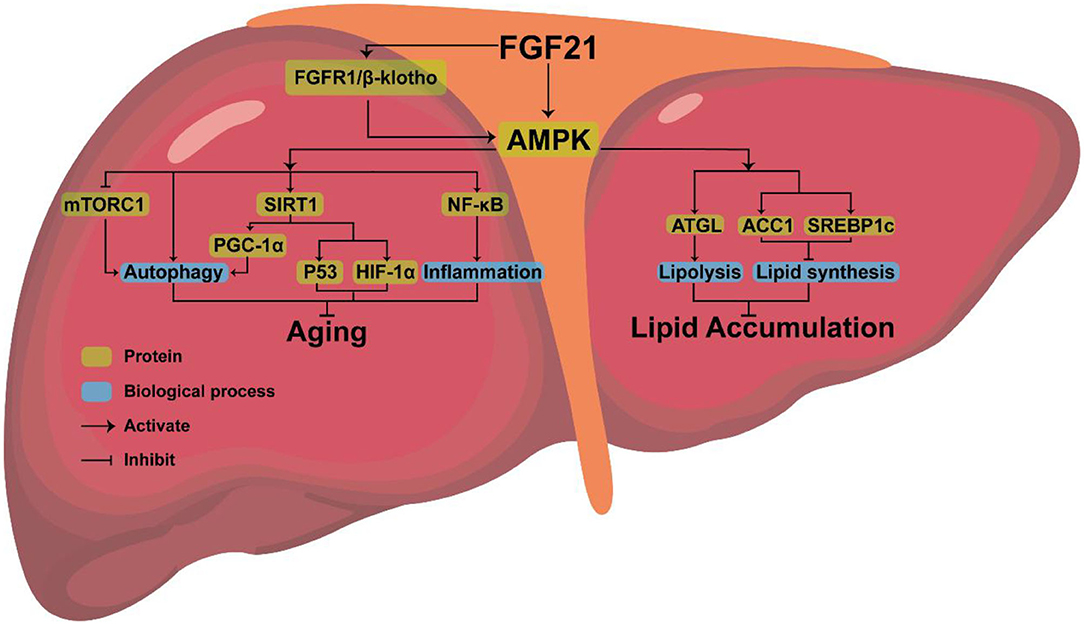
FGF21, a new type of endocrine hormone, is primarily produced by liver (61). Klotho proteins include α-klotho and β-klotho, of which β-klotho is an essential part of the FGF21 receptor complex and necessary for promoting high-affinity binding to its homologous FGF receptor (62). FGF21 signals through a receptor complex composed of fibroblast growth factor receptor 1 (FGFR1) and the coreceptor β-klotho, both required for FGF21 signaling and then activates downstream genes, to exerts its effect (63, 64). Studies have found that, as an endocrine messenger, FGF21 could induce hormonal responses in other tissues, such as the secretion of adiponectin from fat tissue and corticotropin-releasing hormone (CRH) from the hypothalamus, to maintain metabolic homeostasis (65).
In addition, endocrine FGF21 can stimulate the secretion of digestive enzymes from pancreatic acinar cells, which require signaling through a tyrosine kinase receptor complex composed of an FGF receptor and β-Klotho to enhance the digestion of food in stomach (66). FGF21 can also indirectly maintain metabolic homeostasis by activating downstream pathways (67). In cultured adipocytes, FGF21 could regulate metabolism by activating MAPK and downstream ERK1/2, which triggers the activation of GLUT1 and glucose uptake (68). In liver, FGF21 positively controls the PI3K/AKT, insulin-like growth factor 1 (IGF-1), and mTOR pathways to maintain metabolic homeostasis (69).
AMPK, the downstream protein of FGF21, consists of three subunits (AMPKα, AMPKβ, AMPKγ), each with multiple phosphorylation sites, which can regulate lipid metabolism (70). AMPK phosphorylates sterol regulatory element binding protein-1c (SREBP1c) at Ser372, inhibiting the proteolytic cleavage of the precursor SREBP1c to mature SREBP1c, thereby inhibiting steatosis in diet-induced hepatic insulin-resistant mice (71). AMPK can also indirectly inhibit the expression of SREBP1c by reducing mTORC activity, thus decreasing liver lipid content (72) or phosphorylating adipose triglyceride lipase (ATGL), to stimulate TG lipase activity and activate lipolysis (73).
Amazingly, AMPK is highly conserved in eukaryotes, giving them the ability to expand their lifespan (74). By activating AMPK, FGF21 may delay aging and extend mammals’ lifespan (75). Increased longevity has been observed in transgenic worms expressing the modified AMPK-γ subunit (76, 77). Overexpression of a single AMPK-α subunit in the fat body also extended the life span of fruit flies (78).
As a pro-longevity kinase, AMPK can also prevent cellular aging by activating downstream pathways (79–81). AMPK is associated with some downstream pathways involved in controlling aging, such as rapamycin complex 1 (mTORC1), nuclear factor kappa-B (NF-κB), and sirtuin-1(SIRT1). Low levels of inflammation promote aging (82). AMPK may extend longevity by inhibiting the NF-κB pathway and NF-κB-mediated inflammatory response (83). Activation of mTORC1 will inhibit autophagy and accelerate aging, while FGF21 exposure can inhibit the activation of mTORC1 in the liver resulting in anti-aging effects (84).
AMPK can directly activate SIRT1 (75, 85, 86). After being triggered, SITR3 will mediate autophagy to anti-aging (87). FGF21 regulates mitochondrial biogenesis by activating PGC-1α through the FGF21-AMPK-SIRT1 pathway (88). FGF21 also stimulated the expression of PGC-1α in mouse liver (89) and human dopaminergic neurons (90, 91). Improved mitochondrial function and activation of PGC-1α play a crucial anti-aging role (92). AMPK-SIRT1 axis is also connected to several other aging-linked targets, such as p53 and HIF1α (93). Therefore, it is clear that anti-aging effects can be achieved by activating AMPK, but whether FGF21 can inhibit aging by upregulating the expression of AMPK requires further experimental proof.
Combined with the above analysis, we believe that AMPK, the downstream protein of FGF21, may be the key to FGF21 simultaneous aging and metabolism regulation (Figure 2).
FIGURE 2
reference link :https://www.frontiersin.org/articles/10.3389/fcvm.2021.655575/full
More information: Matthew J. Potthoff, FGF21 suppresses alcohol consumption through an amygdalo-striatal circuit, Cell Metabolism (2022). DOI: 10.1016/j.cmet.2021.12.024. www.cell.com/cell-metabolism/f … 1550-4131(21)00690-2



{kind=link}