









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



It’s known that hair follicles assist in the development of a pimple, but new research suggests the skin cells outside of these hair follicles play a larger role. The findings published in the Feb. 16, 2022 issue of Science Translational Medicine.
“These findings may transform the way we treat acne,” said Richard Gallo, MD, Ph.D., Ima Gigli Distinguished Professor of Dermatology and chair of the Department of Dermatology at UC San Diego School of Medicine.
“Previously, it was thought that hair follicles were most important for acne to develop. In this study, we looked at the cells outside of the hair follicle and found they had a major effect on controlling bacteria and the development of acne.”
To counter an infection within a hair follicle, the surrounding skin undergoes a process called reactive adipogenesis in which fibroblasts transform into fat cells. Cathelicidin is produced as well to help combat the infection by suppressing bacteria that can cause acne.
The discovery of cathelicidin’s role came as a surprise.
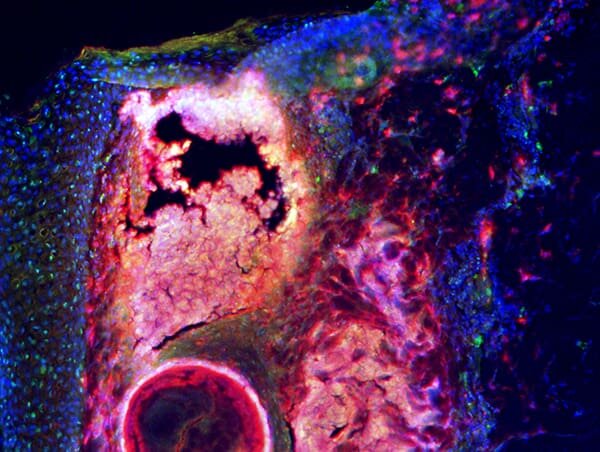
“We began our research wanting to understand the biology of acne and specifically looked at the role of fibroblasts, which typically provide structural support in the deeper layers of the skin,” said first author Alan O’Neill, Ph.D., project scientist at UC San Diego School of Medicine.
“What we uncovered instead was that these cells were activated to produce large amounts of an important antimicrobial, cathelicidin, in response to acne-causing bacteria called Cutibacterium acnes.”
The research team performed skin biopsies on acne patients treated for several months with retinoids, a class of chemicals derived from vitamin A found to promote skin health. To the researchers’ surprise, the drug enhanced cathelicidin expression after treatment, thus finding an additional, unknown mechanism for why retinoids help treat acne.
“Cathelicidin being so highly expressed in acne biopsy tissue was a very interesting finding to us,” said Gallo. “Knowing this will be helpful in developing a more targeted therapy to treat acne.”
Currently, retinoid treatment focuses on controlling the development of lipids in skin cells. One major side effect of these drugs are their teratogenic effects, causing fetal abnormalities in pregnant people. This limits the use of these drugs to only severe cases. The research team hopes these findings may assist in developing a more targeted approach to treating acne.
“This research could assist in identifying new treatment options that specifically target the fibroblast’s ability to produce cathelicidin,” said O’Neill. “Thus creating a therapeutic for acne that would be more selective with potentially less harmful side effects.”
Biological Role of Cathelicidins
LL-37 and mCRAMP
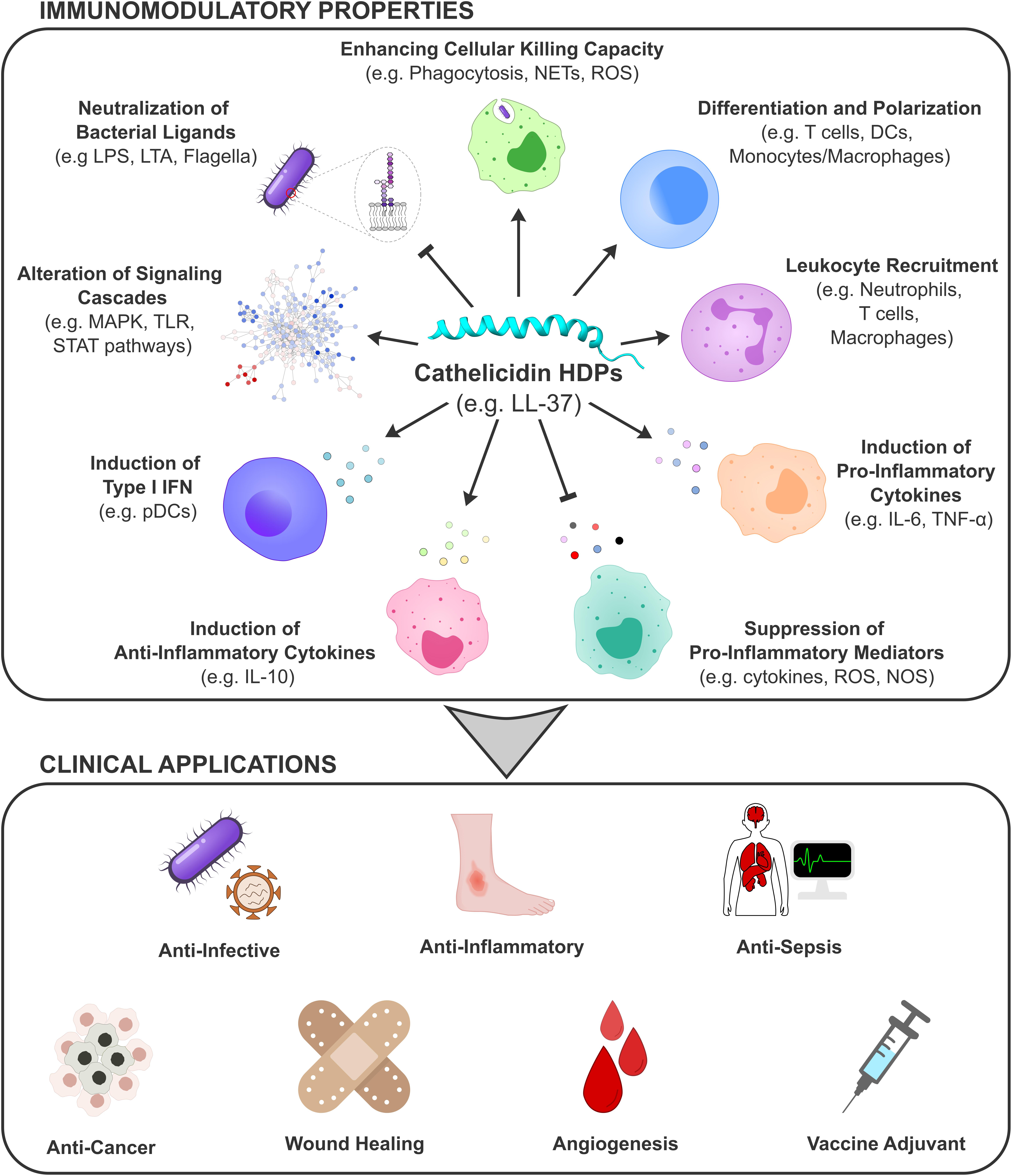
HDPs, in general, exert an incredible array of immunomodulatory functions and many of these features are shared by members of the cathelicidin family of peptides (Figure 2), although individual peptides tend to favor a subset of these properties (reviewed in ref 4). Of the many biological functions of HDPs, their antimicrobial functions have undoubtedly been the most widely researched in part due to the simple assays involved.
While many studies have emphasized the important role of cathelicidins as antimicrobials at epithelial surfaces, particularly the skin (Travis et al., 2000), such conclusions must be qualified due to the conditions under which such activities were assessed, often in very dilute salts. For instance, phosphate buffer, in which many of these studies were undertaken, is decidedly not physiological since in vivo conditions involve high concentrations of divalent and monovalent cations and polyanionic sugars that can inhibit antimicrobial activity (Bowdish et al., 2005b).
Admittedly, studies in mice deficient in the cathelin-related antimicrobial peptide (mCRAMP), the murine ortholog of LL-37, have demonstrated an enhanced susceptibility to a variety of infections including necrotizing skin infections caused by Group A Streptococcus (Nizet et al., 2001), keratitis produced by P. aeruginosa (Huang et al., 2007), and meningitis induced by Streptococcus pneumoniae (Merres et al., 2014).
However, it is possible that such studies could reflect the immunomodulatory effects of cathelicidins that enables protection against infections (Bowdish et al., 2005b; Hancock et al., 2016). In the following section, we discuss studies demonstrating that cathelicidins, such as LL-37, have a primary role in modulating the (innate) immune response which is robust, complex, and occurs under physiological conditions both in vitro and in animal models.
One outstanding feature of LL-37 is its ability to suppress pro-inflammatory signaling. This likely involves a complex series of both direct and indirect mechanisms (Koo and Seo, 2019). Regarding direct mechanisms, cathelicidins bind to and neutralize the bacterial toll-like receptor (TLR) ligands, such as lipopolysaccharide (LPS) or lipoteichoic acid (LTA) (Kandler et al., 2006), which would otherwise engage TLRs and trigger inflammatory processes associated with cascading activation of immune cells (Horibe et al., 2013).
In addition, LL-37 can substantially attenuate LPS-mediated TNF-α production from peripheral blood mononuclear cells (PBMCs) when added either before or after the LPS stimulus, consistent with the notion of a variety of indirect mechanisms that reflect LL-37 uptake into cells and modulation of various intracellular signaling events (Hilchie et al., 2013).
Indeed, in PBMCs challenged with TLR-2, TLR-4, or TLR-9 agonists, LL-37 generally suppresses the production of the pro-inflammatory cytokines TNF-α and IL-1β and alters expression of IL-6 and IL-8 (Mookherjee et al., 2006; Hilchie et al., 2013). Moreover, based on microarray analysis, more than 160 genes up-regulated in LPS-stimulated THP-1 monocytic cells were suppressed in LL-37 treated cells (Mookherjee et al., 2006). Thus, LL-37 neutralizes bacterial signature molecules (TLR agonists) that normally induce pro-inflammatory immune responses.
Several lines of evidence support the host-cell-directed activity of cathelicidins that is independent of bacterial ligand binding. For example, LL-37 modulates more than a dozen signaling pathways including the p38, Erk1/2, JNK MAP-kinases, NFκB, PI3K/Akt, Src family kinase, TRIF-IRF, TREM, Wnt/β-Catenin, JAK-STAT, and autophagy signaling pathways independent of LPS, LTA and/or flagellin stimulation (Scott et al., 2002; Mookherjee et al., 2009; Hancock et al., 2016).
Furthermore, systems biology and biochemical studies on human CD14+ monocytes treated with LL-37 showed nearly 500 genes changing expression, reflecting the involvement of many of these pathways in chemokine induction in response to LL-37 (Mookherjee et al., 2009).
Activation of Wnt/β-Catenin and PI3K/Akt signaling cascades elicited by LL-37 was also demonstrated in samples from non-small cell lung carcinoma patients (Ji et al., 2019) with a positive correlation between LL-37 concentration in tissues and relevant gene expression.
Similarly, in the A549 pneumocyte cell line in vitro, LL-37 stimulated growth through a β-Catenin-dependent but TLR-independent manner (Ji et al., 2019). Overall, such studies provide evidence that LL-37 has multiple surface and intra-cellular targets (Hilchie et al., 2013; Levast et al., 2019) that contribute to the biological functions of this HDP in vivo, resulting in diverse outcomes mediated by a broad array of signaling events (Figure 2).
A typical outcome is the induction of certain chemokines in a variety of cell types, including phagocytes and epithelial cells. Although the resting concentrations of LL-37 at mucosal surfaces can be quite low, local LL-37 concentrations at sites of infection or during acute inflammation can be much higher due to degranulation of phagocytes releasing LL-37 into the vicinity.
The high local concentration promotes the well-studied ability of cathelicidins to enhance the recruitment of immune cells, especially phagocytes (monocytes, macrophages, and neutrophils), to the sites of infections in mice (Hilchie et al., 2013). For example, the release of LL-37 by human keratinocytes activate the Src family kinases and enhance TLR-5 activation upon flagellin stimulation, thereby inducing the release of chemokines (Nijnik et al., 2012).
LL-37 and other HDPs can also act as chemokines to directly attract immune cells. Thus, binding of the FPR2 receptor by LL-37 enables the recruitment of neutrophils, monocytes, T cells, and mast cells (Yang et al., 2000) although, generally speaking, this occurs at higher peptide concentrations than other chemokine inducing activities, likely due to low ligand binding affinity.
However, not all of the functions of LL-37 are beneficial. For example, LL-37 promotes histamine release from mast cells, which promotes loosening of blood vessel walls to enable enhanced uptake of immune cells, but this process is also allergenic (Niyonsaba et al., 2002).
Mast cell degranulation (and histamine release) is mediated by LL-37-induced activation of the MrgX2 receptor that triggers a range of signaling pathways such as PI3K/Akt, Erk1/2, and JNK (Yu et al., 2017). LL-37 also promotes microbially-induced apoptosis of epithelial cells while extending the lifetime of neutrophils (Barlow et al., 2010), which could contribute to damage associated with respiratory infections.
TLRs play a crucial role in innate immunity and govern pathogen recognition by and activation of important sentinel cells, such as macrophages, neutrophils, dendritic cells (DCs), and epithelial cells (Beutler, 2009). In addition to their ability to suppress proinflammatory signaling through TLRs, cathelicidins can also influence the expression of TLRs in a variety of cell types including mast cells, monocytes, neutrophils, renal cells, epithelia lining the colon, and other mucosal surfaces (Agier et al., 2018).
Interestingly, the influence of LL-37 on TLR expression is both tissue- and time- dependent. For example, protein levels of TLR-2, TLR-4, TLR-5, and TLR-9 increased in Wistar rat mast cells in the presence of LL-37 in a time-dependent manner and peaked 3 hours following stimulation in vitro (Agier et al., 2018).
Furthermore, LL-37 downregulated TLR and co-receptor expression induced by LPS in human gingival fibroblasts, but did not influence expression in untreated cells (Inomata et al., 2019), although the relevance of this is unknown since, as mentioned above, LL-37 strongly reduces LPS/LTA-mediated proinflammatory cytokine expression (Kandler et al., 2006).
Cathelicidins can alter the host immune response prior to, during, and even after infection. By directly influencing lymphocytes while also altering the chemokine profile associated with T and B cells, as well as affecting innate immune responses (e.g., cytokines, etc.) that prime and activate lymphocytes, cathelicidins link the innate and adaptive immune systems (Mookherjee et al., 2009; Nicholls et al., 2010).
LL-37 and its derivatives also have the ability to induce differentiation and maturation of DCs as well as activate plasmacytoid DCs (pDC) and macrophages that prime and activate adaptive immunity (Davidson et al., 2004; Kandler et al., 2006). Overall, the features of LL-37 that promote modulation of the immune system, rather than direct microbicidal effects, seem to assist the resolution of an infection while also regulating harmful inflammation.
The immunomodulatory effects of cathelicidins include selectively enhancing and diminishing inflammation by direct and/or indirect chemotaxis, pro-inflammatory and anti-inflammatory cytokine production, blocking TLR activation and downstream signaling pathways, and promoting activation of adaptive immunity.
reference link :https://www.frontiersin.org/articles/10.3389/fmicb.2020.01902/full
More information: Alan M. O’Neill et al, Antimicrobial production by perifollicular dermal preadipocytes is essential to the pathophysiology of acne, Science Translational Medicine (2022). DOI: 10.1126/scitranslmed.abh1478



{kind=link}