









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



I risultati dello studio sono stati pubblicati sulla rivista peer review: Cancers.
https://www.mdpi.com/2072-6694/13/11/2676/htm
I pazienti oncologici sono tradizionalmente considerati ad alto rischio di infezioni virali respiratorie a causa della loro sottostante immunosoppressione [11]. Le infezioni respiratorie hanno un carico particolarmente letale nei pazienti oncologici, data la loro ridotta capacità di combattere le infezioni a causa dell’alterata risposta immunitaria [11].
Ciò porta alla domanda, i pazienti oncologici hanno maggiori probabilità di sviluppare complicazioni letali dopo essere stati infettati da SARS-CoV-2? I primi studi che studiano la relazione tra SARS-CoV-2 e cancro hanno mostrato tassi di mortalità più elevati per infezione da SARS-CoV-2 nei pazienti oncologici rispetto agli individui senza cancro [14].
Sebbene nell’ultimo anno sia stato compiuto un enorme sforzo per raccogliere dati su COVID-19 e cancro, i risultati riportati dovrebbero essere interpretati con cautela. In particolare, i pazienti con un trattamento in corso o recente per tumori ematologici, polmonari o metastatici sono a maggior rischio di sviluppare COVID-19 grave [14,15]. Non è chiaro quanto dell’aumento dei tassi di mortalità sia dovuto al ritardo nella diagnosi e alla gestione non ottimale del cancro a causa della pandemia [15].
La stessa disregolazione immunitaria causata da SARS-CoV-2 potrebbe fornire complicazioni ancora più significative per questa sottopopolazione già vulnerabile. Meccanicamente, l’interazione tra l’ambiente immunitario dell’ospite e il cancro o le infezioni da SARS-CoV-2 utilizza percorsi simili, come ipercoagulabilità, risposta immunitaria disregolata, livelli elevati di citochine, espressione alterata ACE-2 e TMPRSS2 e stato protrombotico, che portano l’essere umano corpo in grave squilibrio e può aggravare gli effetti di SARS-CoV-2 in alcuni malati di cancro [15]. Una guida specifica per quanto riguarda il trattamento del cancro nei pazienti COVID-19 è carente a causa della conoscenza minima, in particolare per quanto riguarda le risposte immunitarie differenziali.
È stato anche dimostrato che le cellule epiteliali del colon esprimono alti livelli di ACE2 e possono supportare la replicazione virale di SARS-CoV-2, con conseguente disfunzione della barriera intestinale [19]. Inoltre, sono presenti le proteasi dell’ospite accessorie necessarie per l’elaborazione dello spike e l’esposizione al dominio S2, inclusi TMPRSS2 e TMPRSS4 [5]. L’infiammazione locale risultante negli epiteli intestinali in combinazione con l’infiammazione sistemica dall’infezione respiratoria da SARS-CoV-2 può avere un impatto significativo sul microbiota intestinale residente.
In precedenza è stato dimostrato che l’infezione influenzale altera significativamente la composizione del microbiota intestinale [20]. Gli interferoni prodotti nei polmoni portano all’esaurimento dei batteri anaerobici obbligati e all’arricchimento dei proteobatteri nell’intestino, portando alla disbiosi. L’azione di questi interferoni ha dimostrato di inibire le risposte antimicrobiche e infiammatorie nell’intestino durante la colite indotta da Salmonella, che ha dimostrato di migliorare ulteriormente la colonizzazione e la disseminazione intestinale della Salmonella, un fattore di rischio per il cancro del colon [21].
Inoltre, i pazienti con sindrome dell’intestino irritabile (IBD) avevano un rischio maggiore di influenza e l’infiammazione IBD ha avuto un impatto sulla risoluzione dei sintomi simil-influenzali [22]. Ciò è significativo dato che l’IBD è un importante fattore di rischio per il CRC ed è anche noto che il microbiota intestinale all’interno di questa popolazione è in genere altamente disbiotico. È noto che la disbiosi intestinale può influenzare la carcinogenesi e la progressione del CRC [23,24].
Inoltre, un altro studio ha riportato la remissione transitoria del linfoma refrattario a cellule NK/T durante l’infezione da COVID-19 e la ricaduta dopo la risoluzione di COVID-19 in un singolo paziente [27]. Questi rapporti insieme suggeriscono che le risposte immunitarie SARS-CoV-2 hanno dimostrato di sopprimere la citotossicità delle cellule NK [28].
È stato anche ipotizzato che l’immunità antivirale delle cellule T CD8+ indotta dalle infezioni da SARS-CoV-2 potrebbe essere utilizzata per migliorare le immunoterapie contro il cancro [29]. Sebbene questi studi forniscano la premessa fondamentale per l’impatto di SARS-CoV-2, un’ulteriore comprensione di come le infezioni virali respiratorie, in particolare SARS-CoV-2, influenzino la progressione del CRC rappresenta un divario di conoscenze e un’esigenza critica insoddisfatta.
Attualmente, si sa poco sull’effetto che SARS-CoV-2 avrà sul microbioma ospite, per non parlare di ambienti di nicchia, come il microbiota intestinale e la sua relazione con il CRC. Secondo l’American Cancer Society, si stima che il CRC abbia causato oltre 140.000 nuovi casi e 50.000 morti negli Stati Uniti nel solo 2020. Una comprensione dettagliata di come l’infezione da SARS-CoV-2 influisca sul microbiota intestinale e sulla progressione del CRC è della massima importanza [30].
Il microbioma intestinale e il CRC
La morte del CRC rimane una delle principali mortalità correlate al cancro negli Stati Uniti, ma il meccanismo molecolare del suo sviluppo non è completamente compreso. Gli studi hanno scoperto che negli Stati Uniti i tassi di incidenza del CRC sono aumentati del 2% all’anno tra gli adulti di età inferiore ai 55 anni dal 2007 al 2016 [30], il che ha portato alla raccomandazione di abbassare l’età per iniziare lo screening colonscopico per il CRC.
Nell’ultimo decennio, l’iniziativa del Progetto Microbioma Umano del National Institutes of Health ha messo in evidenza l’importanza del microbiota intestinale per la salute umana. Il microbiota intestinale svolge un ruolo cruciale nella salute attraverso le sue attività protettive, trofiche e metaboliche. Il microbiota intestinale è stato recentemente collegato allo sviluppo di CRC, poiché è stato dimostrato che i metaboliti derivati dal microbiota influenzano direttamente la cancerogenesi [31].
I microbi anaerobici fermentano i componenti alimentari non digeriti che raggiungono l’intestino crasso per produrre un’ampia gamma di metaboliti, con i prodotti di fermentazione significativi negli adulti sani sono gas e acidi organici: acidi grassi a catena corta (SCFA), come acido acetico, acido propionico , e acido butirrico e acidi grassi a catena media (MCFA), come acido linoleico, acido laurico e acido oleico [32].
Vengono quindi rilasciati nel lume del colon e funzionano come molecole di segnalazione tra i batteri e l’ospite [31]. È stato anche dimostrato che gli SCFA mantengono la barriera intestinale e producono un effetto antinfiammatorio introducendo la differenziazione delle cellule Treg e l’espressione di citochine antinfiammatorie [33]. È importante sottolineare che queste attività nell’intestino formano assi multi-organo, come “l’asse intestino-polmone” e “l’asse intestino-cervello”,
La disbiosi intestinale consente ai metabolomi e ai prodotti batterici di entrare nel sistema circolatorio, portando a un’infiammazione sistemica [36]. L’analisi fecale identifica una marcata disbiosi nei pazienti con CRC provenienti da coorti sane [23]. La disbiosi del microbiota intestinale può influenzare lo stato immunitario, la carcinogenesi e la progressione del tumore nel CRC [37,38]. Gli studi sul CRC hanno dimostrato alterazioni continue nel microbiota intestinale durante lo sviluppo del tumore e queste alterazioni sono direttamente responsabili della progressione del tumore [39]. In uno studio, la colonizzazione di topi privi di germi con microbiota da topi portatori di tumore ha aumentato significativamente la tumorigenesi [40].
I risultati tipici nei pazienti con CRC sono una marcata diminuzione di Firmicutes, Bacteroides e Actinobacteria, abbinata ad un aumento delle popolazioni di Fusobacterium e Porphyromonas [41]. Gli actinobatteri sono uno dei quattro principali phyla del microbiota intestinale. Sebbene rappresentino solo una piccola percentuale della comunità batterica, gli actinobatteri sono fondamentali nel mantenimento dell’omeostasi intestinale e della tolleranza immunitaria [42]. Popolazioni sbilanciate di Actinobacteria sono state evidenziate in diverse condizioni patologiche [43].
In particolare per il CRC, la riduzione dei metaboliti associati al phylum degli Actinobacteria accoppiata con una risposta immunitaria proinfiammatoria, cellulo-mediata, citotossica delle cellule T helper-1 (Th1) indotta da questo gruppo si presta a una prognosi sfavorevole del CRC [44]. Una diminuzione dei batteri produttori di butirrato (in particolare Actinobacteria e Firmicutes phyla) diminuisce anche la fonte di energia primaria per i colonociti.
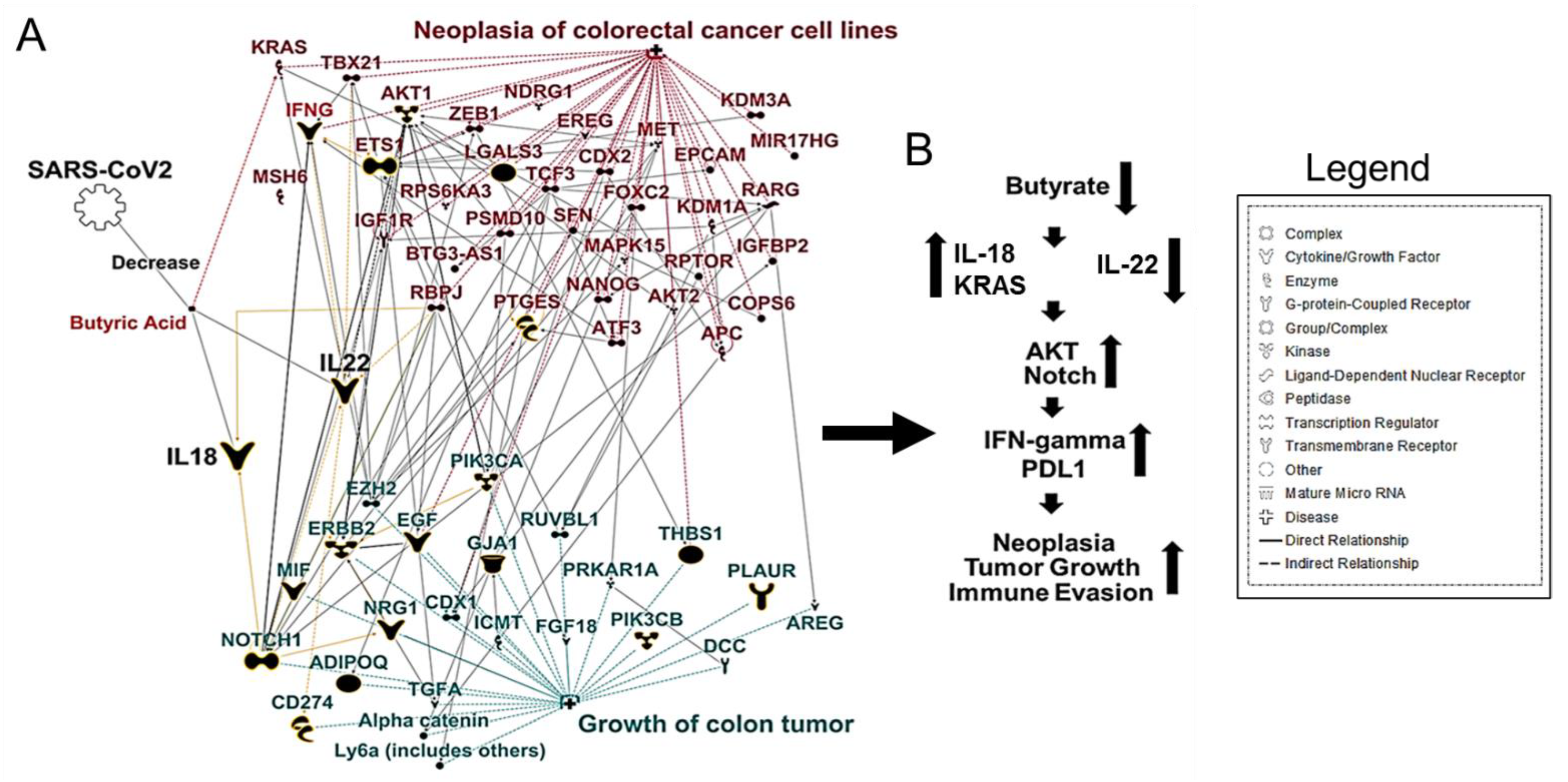
Aumenta il pH intracolonico, creando un ambiente ostile per i colonociti e contribuendo alla tumorigenesi [41]. Per esplorare ulteriormente la relazione tra butirrato e CRC, abbiamo utilizzato Ingenuity Pathway Analysis (IPA) [45] che utilizza un database di conoscenze biologiche precedenti derivate dall’analisi e dall’annotazione della letteratura scientifica sottoposta a revisione paritaria.
Abbiamo utilizzato gli strumenti My Pathway/Path Designer in IPA per tracciare le interazioni note tra butirrato e CRC. Abbiamo scoperto che la disbiosi intestinale potrebbe causare una diminuzione dei batteri produttori di butirrato. Una successiva mancanza di produzione di IL-22 consente ai metaboliti batterici di entrare nel sistema circolatorio, portando a una maggiore infiltrazione di cellule infiammatorie, aumentando così la colite associata al cancro del colon (Figura 1A, B).
Il consumo di una dieta ricca di grassi e proteine è stato collegato a una maggiore secrezione di acidi biliari. Alcuni microbi si convertono in acidi biliari primari e secondari, che sono tossici e promuovono la tumorigenesi [46]. Gli acidi biliari sono molecole di segnalazione chiave che regolano le funzioni digestive e le funzioni fisiologiche come il metabolismo del glucosio e dei lipidi e l’omeostasi immunitaria [47].
Gli acidi biliari sono sintetizzati dal colesterolo e coniugati con glicina o taurina negli epatociti. La maggior parte degli acidi biliari viene secreta nell’intestino tenue. Tuttavia, gli acidi biliari che raggiungono l’intestino crasso interagiranno con i microbi intestinali. Gli acidi biliari sono tossici per molti batteri intestinali. Pertanto, livelli elevati eserciteranno una pressione di crescita selettiva sul nostro microbiota intestinale, favorendo alcuni batteri nel nostro intestino ad agire enzimaticamente sugli acidi biliari [47].
Ad esempio, la taurina in eccesso viene escreta come acido biliare coniugato chiamato acido taurocolico e convertito in acido desossicolico, che studi hanno dimostrato di essere genotossico e promuovere la formazione di tumori [48].
Negli studi, l’acido desossicolico può attivare vie di segnalazione cellulare associate alla proliferazione cellulare e all’apoptosi [46]. Un importante enzima nella produzione di acido desossicolico, la 7α-deidrossilazione, è stato caratterizzato in specie appartenenti ai generi Eubacterium e Clostridium, comprese le specie Clostridium scindens e Clostridium hylemonae [47].
È stato dimostrato nei topi che aumenta la proporzione di Lactobacillus e Bifidobacterium nel microbiota intestinale attraverso l’integrazione di probiotici (gavage orale al giorno di 0,6 miliardi di UFC (unità formanti colonie) ciascuno di Lactobacillus acidophilus, Lactobacillus rhamnosus e Bifidobacterium bifidum, diluito in 200 μL di acqua potabile) ridotta infiltrazione di cellule infiammatorie, ridotta espressione di chemochine e ridotto CRC associato alla colite [49].
Altri studi hanno suggerito che un’espansione dei Proteobacteria, solitamente un costituente minore del microbiota intestinale, è una potenziale firma microbica della disfunzione epiteliale osservata nei pazienti con CRC [39,50,51]. Una comprensione completa dei complessi fattori che portano alla disbiosi non si è ancora materializzata e l’effetto di ciò sul CRC rimane poco chiaro.
Disbiosi e CRC del microbioma intestinale indotto da SARS-CoV-2
Sebbene i sintomi COVID-19 più comuni siano respiratori, le infezioni da SARS-CoV-2 colpiscono anche il tratto gastrointestinale, culminando in infiammazione e crampi intestinali fino alla diarrea [52]. Un’ipotesi è che la downregulation di ACE2, causata dall’infezione da SARS-CoV-2, riduca l’attivazione del target meccanicistico della rapamicina (mTOR) e aumenti l’autofagia, portando a disbiosi intestinale e diarrea [52]. L’espressione di ACE2 nei polmoni è sottoregolata nei topi wild-type infettati da SARS-CoV e nei topi ai quali è stata iniettata la proteina spike ricombinante della SARS. Questa downregulation può svolgere un ruolo nella patogenesi della SARS e nella progressione della malattia verso l’ARDS [53,54,55].
Un’altra ipotesi è che l’intestino tenue sia probabilmente un sito chiave di amplificazione della risposta infiammatoria sistemica, dove il blocco dell’ACE2 provoca un aumento dei livelli di angiotensinogeno e un’iperattivazione del sistema renina-angiotensina, portando all’arresto del trasportatore di aminoacidi BA0T1 e a un conseguente mancanza di triptofano cellulare, che porta a una ridotta secrezione di peptidi antimicrobici e disbiosi intestinale [56].
Studi recenti hanno osservato che i pazienti con COVID-19 sviluppano un microbiota disbiotico dell’intestino [24,57,58,59,60,61] (Figura 2). Uno studio ha studiato i cambiamenti nei microbiomi fecali di pazienti con infezione da SARS-CoV-2 durante il ricovero e le associazioni con la gravità e la diffusione fecale del virus [24]. I pazienti con COVID-19 presentavano alterazioni significative nei loro microbiomi fecali, caratterizzati dall’arricchimento di patogeni opportunisti e dall’esaurimento dei commensali benefici al momento del ricovero e in tutti i momenti durante il ricovero.
I simbionti impoveriti e la disbiosi intestinale sono persistiti durante il ricovero, anche dopo l’eliminazione di SARS-CoV-2 e la risoluzione dei sintomi respiratori. Durante il ricovero, Bacteroides dorei, Bacteroides taiotaomicron, Bacteroides massiliensis e Bacteroides ovatus, che sottoregolano l’espressione di ACE2 nell’intestino murino, sono correlati inversamente con il carico SARS-CoV-2 nei campioni fecali dei pazienti. È interessante notare che l’abbondanza di base di microbi specifici nell’intestino, come Coprobacillus, Clostridium ramosum e Clostridium hathewayi, era correlata alla gravità del COVID-19. C’era una correlazione inversa tra l’abbondanza di Faecalibacterium prausnitzii, un batterio antinfiammatorio, e la gravità della malattia.
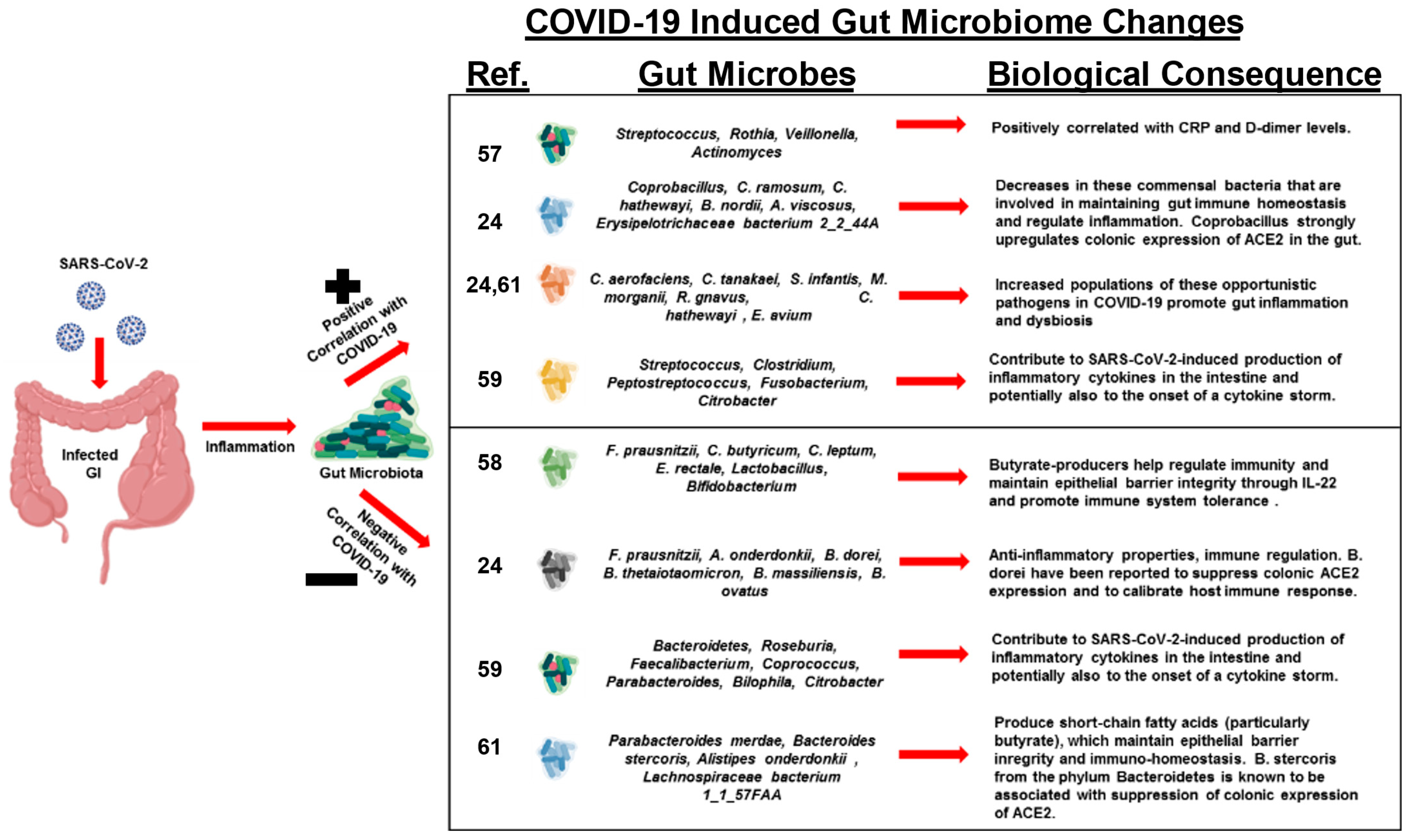
Un altro studio ha studiato i cambiamenti nell’abbondanza di dieci gruppi batterici intestinali predominanti nei pazienti COVID-19 per stabilire una correlazione tra questi gruppi batterici e gli indicatori clinici di polmonite [58]. I risultati indicano che la disbiosi si è verificata nei pazienti con COVID-19 e i cambiamenti nella comunità microbica intestinale erano associati alla gravità della malattia e ai parametri ematologici. È stato dimostrato che il numero di patogeni opportunisti comuni, Enterococcus (Ec) e Enterobacteriaceae (E), è aumentato nei pazienti COVID-19, specialmente nei malati critici. Le osservazioni suggeriscono che questi gruppi batterici possono fungere da biomarcatori diagnostici per COVID-19 e che il rapporto Ec/E può essere utilizzato per predire la morte nei pazienti critici.
Il prossimo studio ha studiato l’attività trascrizionale di SARS-CoV-2 e la sua associazione con le alterazioni del microbioma fecale nei pazienti con COVID-19 [61]. Anche in assenza di sintomi gastrointestinali (GI), alcuni pazienti hanno continuato a mostrare una firma di infezione virale attiva fino a 6 giorni dopo l’eliminazione di SARS-CoV-2 dai campioni respiratori. I campioni fecali con una firma SARS-CoV-2 elevata presentavano abbondanze maggiori delle specie batteriche Collinsella aerofaciens, Collinsella tanakaei, Streptococcus infantis e Morganella morganii. Il microbiota intestinale dei pazienti con infezione attiva da SARS-CoV-2 GI era caratterizzato da arricchimento di patogeni opportunisti, perdita di batteri salutari, aumento della capacità funzionale per la biosintesi di nucleotidi e aminoacidi e metabolismo dei carboidrati. Questo studio fornisce prove per un dormiente,
È stato anche dimostrato che i pazienti COVID-19 hanno una diversità batterica significativamente ridotta, una maggiore abbondanza relativa di patogeni opportunisti, come Streptococcus, Rothia, Veillonella e Actinomyces, e un’abbondanza relativa inferiore di simbionti benefici [57,59]. I livelli di Fusicatenibacter, Romboutsia, Intestinibacter, Actinomyces ed Erysipelatoclostridium hanno mostrato un’elevata precisione per distinguere i pazienti COVID-19, il che suggerisce il potenziale valore del microbiota intestinale come biomarcatore diagnostico e bersaglio terapeutico per COVID-19. La ridotta diversità batterica nell’intestino è stata associata a effetti metabolici acuti ea lungo termine e alla propensione alle malattie.
Nei pazienti con COVID-19, l’abbondanza di batteri produttori di butirrato, come Faecalibacterium prausnitzii, Clostridium butyricum, Clostridium leptum ed Eubacterium rectale, è diminuita in modo significativo e questo cambiamento nella comunità batterica può aiutare a discriminare i pazienti in condizioni critiche dai pazienti che hanno uno stato di malattia più moderato [58]. I campioni fecali di pazienti con infettività SARS-CoV-2 da bassa a nulla avevano un’abbondanza maggiore di batteri produttori di SCFA, Parabacteroides merdae, Bacteroides stercoris, Alistipes onderdonkii e Lachnospiraceae bacterium 1_1_57FAA [61].
È importante sottolineare che i batteri produttori di butirrato sono di fondamentale importanza nel mantenimento dell’integrità della barriera intestinale. Gli SCFA sono stati implicati nella segnalazione di IL-22, che mantiene l’integrità della barriera epiteliale intestinale e polmonare [62]. Il butirrato e altri SCFA regolano l’infiammazione dei macrofagi nell’intestino e promuovono l’effetto Warburg, che vincola metabolicamente le cellule neoplastiche [63].
Il CRC ha una dipendenza metabolica dalla glicolisi anaerobica; quindi, l’ossidazione degli acidi grassi è limitata. Dato questo stato metabolico, il butirrato inizia ad accumularsi nel citoplasma dei colonociti cancerosi, consentendo a questo SCFA di agire come un inibitore dell’HDAC, sensibilizzando i colonociti cancerosi alle vie apoptotiche [64,65]. In particolare, una riduzione dei batteri produttori di butirrato e la disbiosi intestinale hanno contribuito a una minore produzione di IL-22,
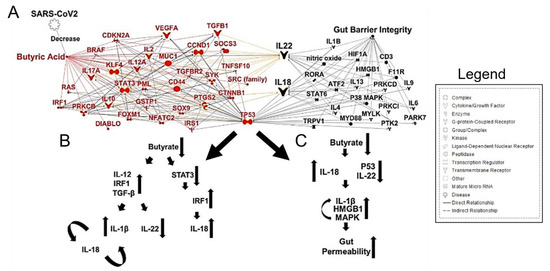
La batteriemia da Fusobacterium nucleatum è stata osservata anche in casi gravi di COVID-19 e ha dimostrato di colonizzare il muco del colon con infiammazione della mucosa associata [67].
Insieme alla disfunzione immunitaria osservata in COVID-19, F. nucleatum provoca immunosoppressione mediante l’azione di una delle sue proteine di superficie, Fap2, che è stata riconosciuta come una lectina legante il galattosio che può interagire con il dominio ITIM delle cellule T (TIGIT) recettore inibitorio [68].
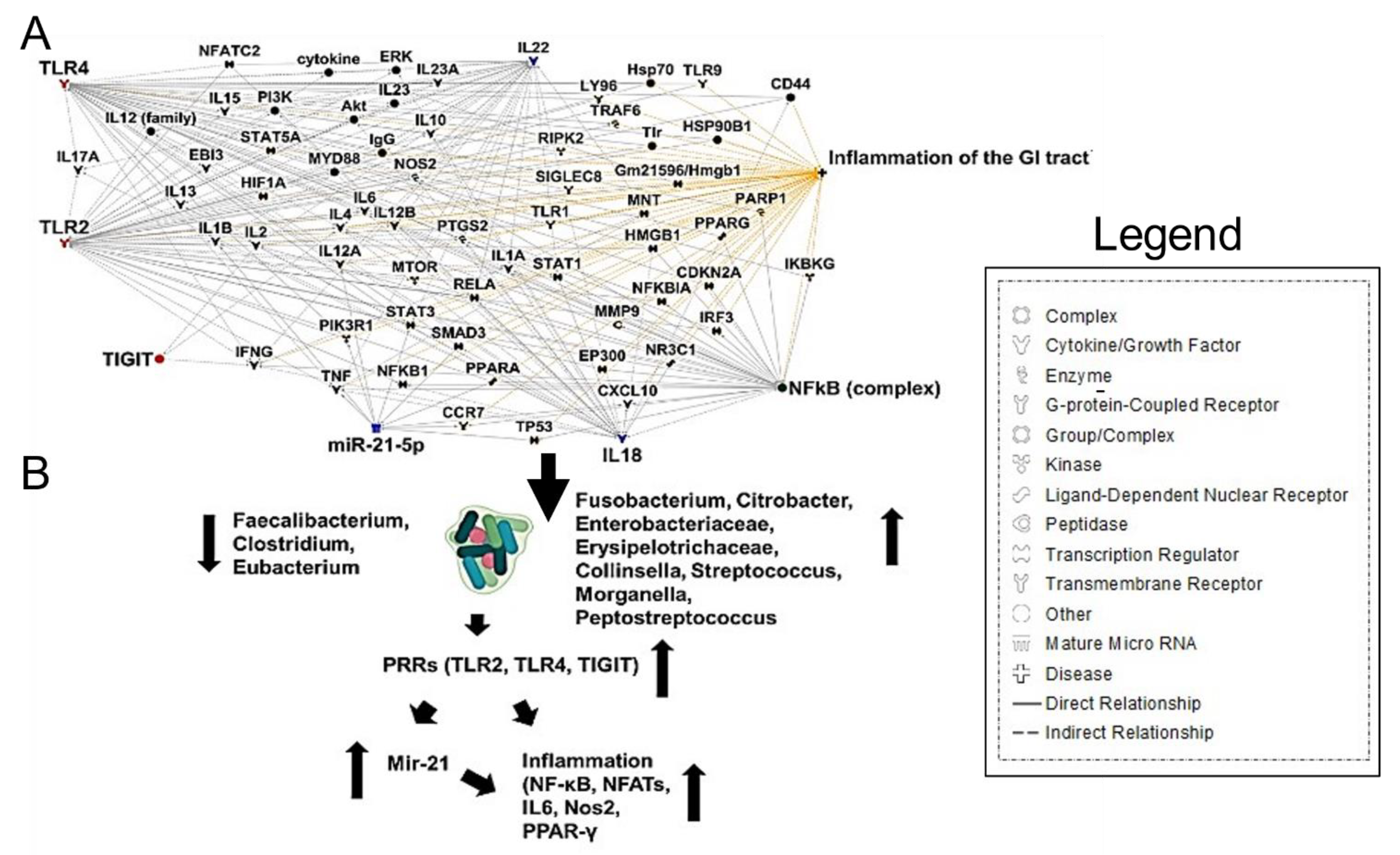
Inoltre, è stato dimostrato che F. nucleatum interagisce con la segnalazione dei recettori Toll-like 2 e 4, con conseguente aumento dell’espressione di microRNA-21 (miRNA21), che promuove l’induzione di NF-kB, contribuendo all’infiammazione e alla proliferazione cellulare [68] (Figura 4A, B). È stato anche dimostrato che la citochina infiammatoria intestinale IL-18 è elevata nel siero dei pazienti COVID-19 [59]. Questi risultati indicano che i cambiamenti della composizione del microbiota intestinale contribuiscono alla produzione di citochine infiammatorie indotta da SARS-CoV-2 nell’intestino.
È interessante notare che uno studio ha anche studiato i cambiamenti nei microbiomi fungini fecali (micobioma) dei pazienti COVID-19 [69]. I pazienti con COVID-19 presentavano alterazioni significative nei loro micobiomi fecali, caratterizzati da proporzioni aumentate di patogeni fungini opportunistici, Candida albicans, Candida auris e Aspergillus flavus. Due agenti patogeni fungini associati alle vie respiratorie, A. flavus e Aspergillus niger, sono stati rilevati in campioni fecali di un sottogruppo di pazienti con COVID-19, anche dopo l’eliminazione della SARS-CoV-2 dai campioni nasofaringei e la risoluzione dei sintomi respiratori.
Questi risultati sono sorprendenti e rafforzano la natura commensale di tutti i microbi, che possono comprendere allo stesso modo batteri, funghi, protisti e virus.
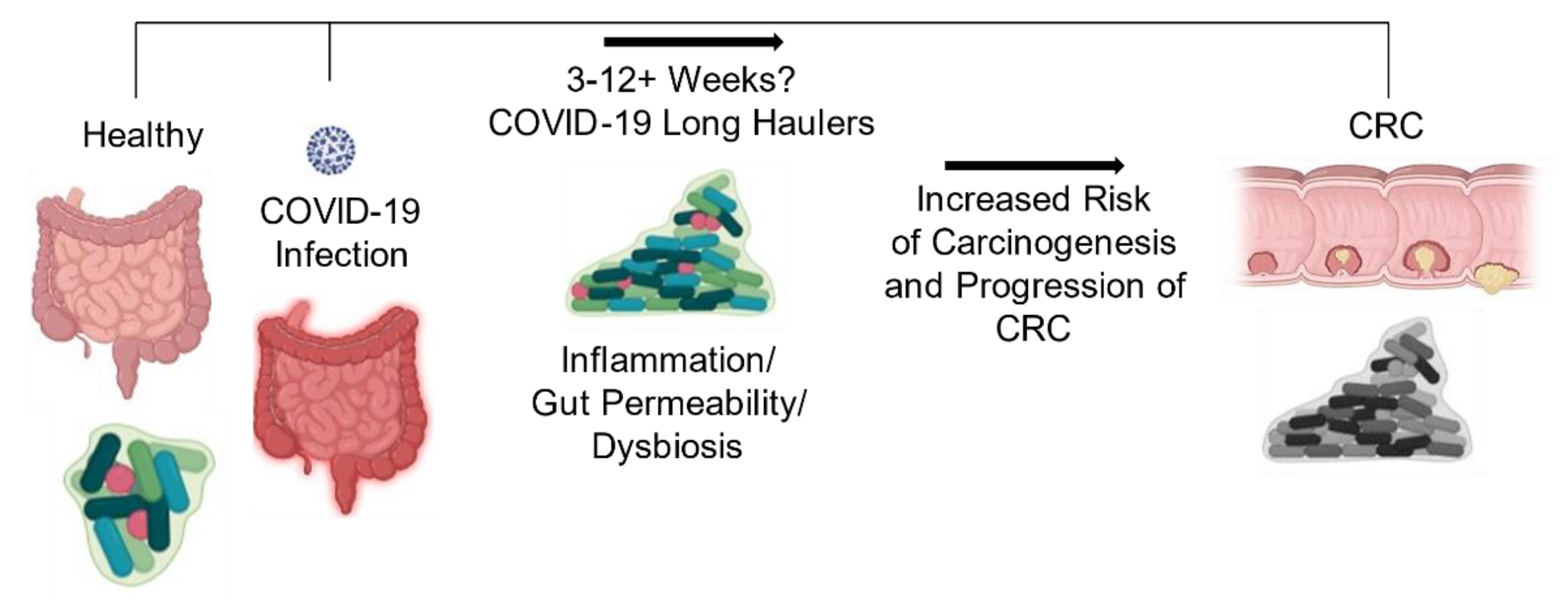
Ci sono segnalazioni crescenti di effetti persistenti e prolungati dopo COVID-19, ora definiti “trasportatori di lunga durata”, una sindrome caratterizzata da sintomi persistenti e complicazioni ritardate o a lungo termine oltre 4-6 settimane dall’insorgenza dei sintomi [70,71]. Ciò può essere causato dalla sindrome da COVID-19 post-acuta osservata osservata in alcuni casi, in cui la disregolazione immunitaria e i sintomi gastrointestinali sono ancora in corso anche dopo l’eliminazione di SARS-CoV-2, influenzando così lo stato del microbioma del paziente [72,73 ]. A causa della natura recente della pandemia di COVID-19, il tempo di ripristino del normale microbiota intestinale deve ancora essere studiato a fondo, quindi la sequenza temporale e gli effetti a lungo termine della disbiosi sono sconosciuti e richiedono ulteriori studi.
Un noto fenomeno di infezione respiratoria è la disfunzione della barriera intestinale, che è correlata a un decorso clinico più grave della malattia [74]. Clinicamente, è stato dimostrato che la modulazione del microbiota intestinale può rallentare la replicazione precoce del virus dell’influenza negli epiteli polmonari, con conseguente riduzione dell’enterite e della polmonite associata al ventilatore [74]. L’espansione delle specie di Bifidobacterium dopo e dopo l’infezione da influenza, ad esempio, ha dimostrato di essere protettiva dall’infiammazione che potrebbe causare eventi disbiotici [75,76]. Attualmente, non ci sono prove cliniche dirette che la modulazione del microbiota intestinale svolga un ruolo terapeutico nel trattamento del COVID-19. Tuttavia, sono in corso studi clinici di probiotici contro COVID-19 [77].
Il sistema immunitario umano si è evoluto per far fronte alla presenza di microrganismi sia all’interno che all’esterno del nostro corpo [78]. Questo fatto significa che la maggior parte delle malattie infettive causate da virus o batteri sono autolimitanti [79,80]. Questa potrebbe essere una possibile ragione per la natura asintomatica o lieve della maggior parte dei casi di COVID-19 [81]. Questi microrganismi possono fungere da fonte di metaboliti, come aminoacidi essenziali e acidi grassi, per il corpo umano [82].
In uno stato di disbiosi intestinale, può verificarsi un transitorio eccesso di apporto nutritivo, poiché sia i microrganismi che il tessuto ospite danneggiato verranno degradati e diventeranno una possibile fonte di nutrimento. Ciò può favorire l’iperinfiammazione durante l’infezione acuta [83] e possibilmente portare a malattie croniche, come il cancro [84].
Ad esempio, parte dell’eccesso di nutrimento da tessuto danneggiato, insieme alla nutrizione in eccesso da un microbioma intestinale disbiotico, potrebbero trasformarsi in intermedi lipidici, causando lipotossicità e ulteriore danno tissutale [85]. Ciò evidenzia come lo stato nutrizionale può influire sulla salute e sull’esito della malattia di una persona. In quanto tale, la restrizione dell’assunzione di cibo potrebbe offrire un modo promettente per controllare l’infiammazione cronica [86].
Insieme, queste osservazioni collettive giustificano una maggiore indagine sulla relazione tra i cambiamenti nel microbiota intestinale che comprendono tutti i microrganismi dovuti all’infezione da COVID-19 e le associazioni con un aumento del rischio di sviluppo o progressione del CRC (Figura 5). Data la diffusa incidenza di questo virus e che il CRC è il terzo tumore più comune diagnosticato negli Stati Uniti sia negli uomini che nelle donne, sono molto necessari studi che indaghino sugli effetti associati di queste due malattie. Dopo aver chiarito le risposte a queste domande, potrebbe essere possibile sviluppare terapie probiotiche efficaci che mantengono un ambiente intestinale più omeostatico durante la malattia, il che potrebbe aiutare a migliorare i tassi di sopravvivenza del CRC.
collegamento di riferimento: https://www.mdpi.com/2072-6694/13/11/2676/htm



{kind=link}