










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



The study findings were published in the peer reviewed journal: Cancers.
https://www.mdpi.com/2072-6694/13/11/2676/htm
Cancer patients are traditionally considered at high risk for respiratory viral infections due to their underlying immunosuppression [11]. Respiratory infections have a particularly lethal burden in cancer patients, given their reduced competency to fight infections due to altered immune responsiveness [11].
This leads to the question, are cancer patients more likely to develop lethal complications after being infected by SARS-CoV-2? Early studies investigating the relationship between SARS-CoV-2 and cancer have shown higher mortality rates from SARS-CoV-2 infection in cancer patients than individuals without cancer [14].
While a massive effort to collect data on COVID-19 and cancer has been performed over the last year, the reported findings should be interpreted with caution. Specifically, patients with ongoing or recent treatment for hematologic, lung, or metastatic cancers are at higher risk of developing severe COVID-19 [14,15]. It is unclear how much of the increase in mortality rates is due to delayed diagnosis and suboptimal cancer management due to the pandemic [15].
SARS-CoV-2-caused immune dysregulation itself could provide even more significant complications for this already vulnerable subpopulation. Mechanistically, the interaction between the host immune environment and cancer or SARS-CoV-2 infections uses similar pathways, such as hypercoagulability, dysregulated immune response, elevated cytokine levels, altered expression ACE-2 and TMPRSS2, and prothrombotic status, which bring the human body into severe disequilibrium and may aggravate the effects of SARS-CoV-2 in some cancer patients [15]. Specific guidance regarding cancer treatment in COVID-19 patients is lacking due to minimal knowledge, particularly regarding differential immune responses.
It has also been shown that colon epithelial cells express high levels of ACE2 and can support viral replication of SARS-CoV-2, resulting in gut barrier dysfunction [19]. Furthermore, the accessory host proteases required for spike processing and S2 domain exposure are present, including TMPRSS2 and TMPRSS4 [5]. The resulting local inflammation in the gut epithelia in conjunction with the systemic inflammation from the respiratory infection from SARS-CoV-2 can significantly impact the resident gut microbiota.
Influenza infection has been previously shown to significantly alter the composition of the intestinal microbiota [20]. Interferons produced in the lungs lead to the depletion of obligate anaerobic bacteria and proteobacteria enrichment in the gut, leading to dysbiosis. The action of these interferons was shown to inhibit the antimicrobial and inflammatory responses in the gut during Salmonella-induced colitis, which was shown to further enhance Salmonella intestinal colonization and dissemination, a risk factor for colon cancer [21].
Furthermore, patients with irritable bowel syndrome (IBD) had an increased risk for influenza, and IBD inflammation impacted the resolution of flu-like symptoms [22]. This is significant given that IBD is a prominent risk factor for CRC, and it is also known that the gut microbiota within this population is typically highly dysbiotic. It is known that dysbiosis of the gut can influence carcinogenesis and the progression of CRC [23,24].
Additionally, another study reported transient remission of refractory NK/T-cell lymphoma during COVID-19 infection and the relapse after COVID-19 resolution in a single patient [27]. These reports together suggest SARS-CoV-2 immune responses have been shown to suppress NK cells cytotoxicity [28].
It has also been hypothesized that the antiviral CD8+ T cell immunity induced by SARS-CoV-2 infections could be used to enhance cancer immunotherapies [29]. While these studies provide the foundational premise for the impact of SARS-CoV-2, further understanding of how respiratory viral infections, particularly SARS-CoV-2, affect CRC progression represents a knowledge gap and critical unmet need.
Currently, little is known about the effect SARS-CoV-2 will have on the host microbiome, let alone on niche environments, such as the gut microbiota and its relation to CRC. According to the American Cancer Society, CRC is estimated to have caused over 140,000 new cases and 50,000 deaths in the United States in 2020 alone. A detailed understanding of how SARS-CoV-2 infection impacts the gut microbiota and CRC progression is of utmost importance [30].
The Gut Microbiome and CRC
CRC death remains one of the leading cancer-related mortalities in the United States, but the molecular mechanism of its development is not fully understood. Studies have found that in the United States, CRC incidence rates have increased by 2% annually among adults younger than age 55 from 2007 to 2016 [30] that has led to the recommendation to lower the age for starting colonoscopic screening for CRC. In the past decade, the Human Microbiome Project’s initiative by the National Institutes of Health has highlighted the importance of the gut microbiota to human health. The gut microbiota plays a crucial role in health through its protective, trophic, and metabolic activities. The gut microbiota has been recently linked to CRC development, as microbiota-derived metabolites have been shown to influence carcinogenesis directly [31].
The anaerobic microbes ferment undigested dietary components that reach the large intestine to produce a wide range of metabolites, with the significant fermentation products in healthy adults being gases and organic acids: short-chain fatty acids (SCFAs), such as acetic acid, propionic acid, and butyric acid and medium-chain fatty acids (MCFAs), such as linoleic acid, lauric acid, and oleic acid [32]. They are then released into the colonic lumen and function as signaling molecules between the bacteria and the host [31]. SCFAs have also been shown to maintain the gut barrier and produce an anti-inflammatory effect by introducing Treg cell differentiation and the expression of anti-inflammatory cytokines [33]. Importantly, these activities in the gut form a multi-organ axes, such as the “gut–lung axis” and the “gut–brain axis”, which are dependent on the metabolic products of the gut in maintaining the healthy homeostatic status of these organ systems [34,35].
Gut dysbiosis allows bacterial metabolomes and products to enter the circulatory system, leading to systemic inflammation [36]. The fecal analysis identifies marked dysbiosis in CRC patients from healthy cohorts [23]. Dysbiosis of the gut microbiota can influence immune status, carcinogenesis, and tumor progression in CRC [37,38]. CRC studies have demonstrated continual alterations in the gut microbiota during tumor development, and these alterations are directly responsible for tumor progression [39]. In one study, the colonization of germ-free mice with microbiota from tumor-bearing mice significantly increased tumorigenesis [40].
Typical findings in CRC patients are a marked decrease in Firmicutes, Bacteroides, and Actinobacteria, paired with increases in Fusobacterium and Porphyromonas populations [41]. Actinobacteria are one of the four major phyla of the gut microbiota. Although they represent only a small percentage of the bacterial community, Actinobacteria are pivotal in maintaining gut homeostasis and immune tolerance [42]. Unbalanced populations of Actinobacteria have been evidenced in several pathological conditions [43].
Specifically for CRC, the reduction in metabolites associated with the Actinobacteria phylum paired with a proinflammatory, cellular-mediated, cytotoxic T-helper cell-1 (Th1) immune response induced by this group lends itself to a poor prognosis of CRC [44]. A decrease in butyrate-producing bacteria (especially the Actinobacteria and Firmicutes phyla) also decreases the primary energy source for colonocytes.
It increases intra-colonic pH, creating a hostile environment for colonocytes and contributing to tumorigenesis [41]. To further explore the relationship between butyrate and CRC, we used Ingenuity Pathway Analysis (IPA) [45] that uses a database of prior biological knowledge derived from analyzing and annotating the peer-reviewed scientific literature. We used the My Pathway/Path Designer tools in IPA to plot known interactions between butyrate and CRC. We found that gut dysbiosis could cause a decrease in butyrate-producing bacteria. A subsequent lack of IL-22 production allows bacterial metabolites to enter the circulatory system, leading to increased inflammatory cell infiltration, thus increasing colitis associated with colon cancer (Figure 1A,B).
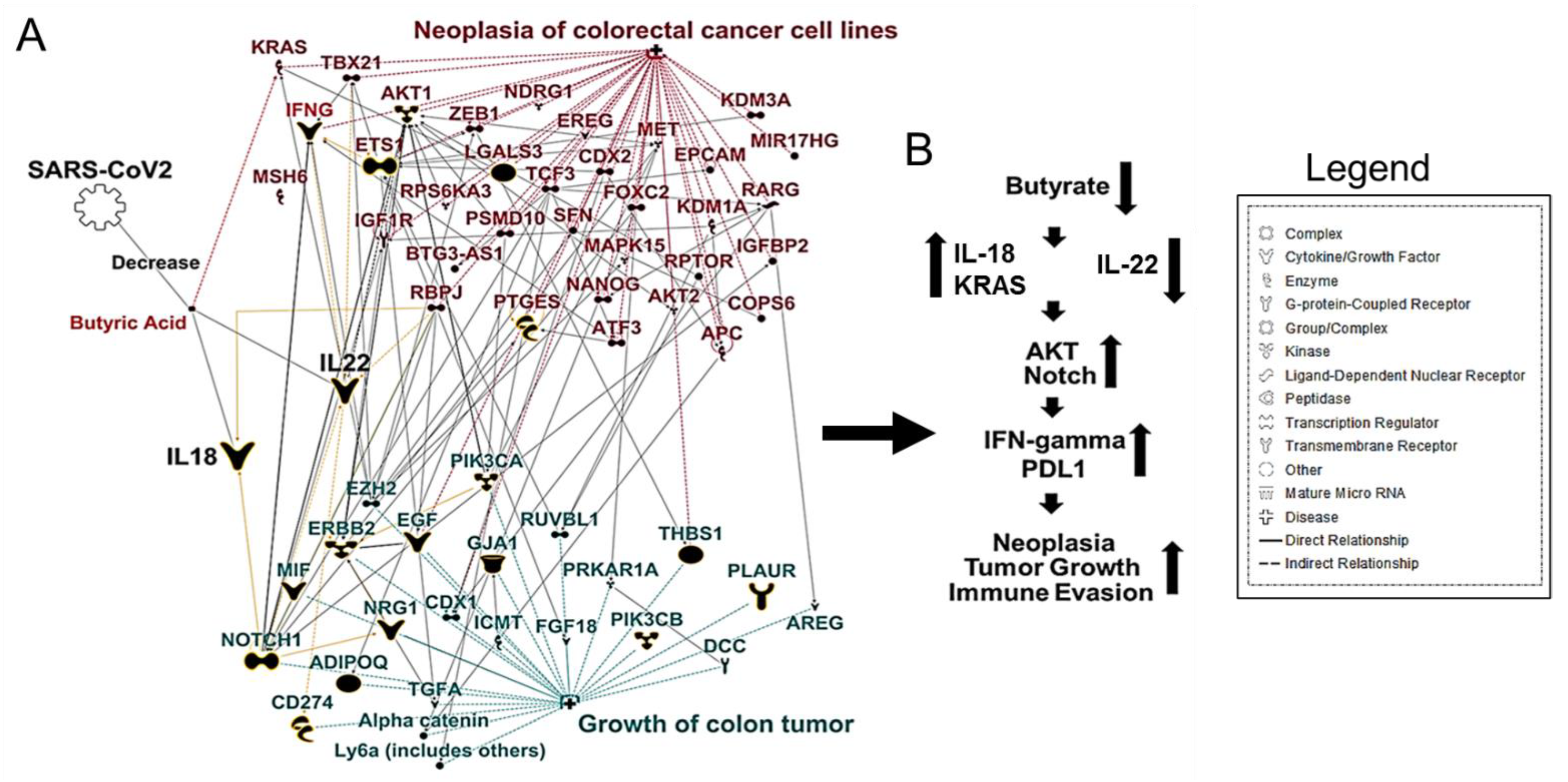
Consumption of a high-fat and -protein diet has been linked to increased secretion of bile acids. Certain microbes convert into primary and secondary bile acids, which are toxic and promote tumorigenesis [46]. Bile acids are key signaling molecules that regulate digestive functions and physiological functions such as glucose and lipid metabolism and immune homeostasis [47].
Bile acids are synthesized from cholesterol and conjugated to glycine or taurine in hepatocytes. Most bile acids are secreted into the small intestine. However, bile acids that reach the large intestine will interact with gut microbes. Bile acids are toxic for many gut bacteria. Thus, high levels will put selective growth pressure on our gut microbiota, favoring certain bacteria in our gut to act enzymatically on the bile acids [47].
For example, excess taurine is excreted as a conjugated bile acid called taurocholic acid and converted into deoxycholic acid, which studies have shown to be genotoxic and promote tumor formation [48].
In studies, deoxycholic acid can activate cellular signaling pathways associated with cell proliferation and apoptosis [46]. A major enzyme in the production of deoxycholic acid, 7α-dehydroxylation, has been characterized in species belonging to the genera Eubacterium and Clostridium, including the species Clostridium scindens and Clostridium hylemonae [47].
It has been shown in mice that increasing the proportion of Lactobacillus and Bifidobacterium in the gut microbiota through probiotic supplementation (oral gavage daily of 0.6 billion CFU (colony forming units) each of Lactobacillus acidophilus, Lactobacillus rhamnosus, and Bifidobacterium bifidum, diluted in 200 μL of drinking water) reduced inflammatory cell infiltration, lowered chemokine expression, and reduced colitis-associated CRC [49].
Other studies have suggested that an expansion of Proteobacteria, usually a minor constituent of the gut microbiota, is a potential microbial signature of epithelial dysfunction observed in patients with CRC [39,50,51]. A complete understanding of the complex factors that lead to dysbiosis has not yet materialized, and the effect of this on CRC remains unclear.
SARS-CoV-2 Induced Gut Microbiome Dysbiosis and CRC
Although the most common COVID-19 symptoms are respiratory, SARS-CoV-2 infections also target the gastrointestinal tract, culminating in inflammation and intestinal cramps to diarrhea [52]. One hypothesis is that ACE2 downregulation, caused by SARS-CoV-2 infection, decreases activation of the mechanistic target of rapamycin (mTOR) and increases autophagy, leading to intestinal dysbiosis and diarrhea [52]. ACE2 expression in lungs is downregulated in wild-type mice infected with SARS-CoV and mice injected with recombinant SARS spike protein. This downregulation may play a role in SARS pathogenesis and disease progression to ARDS [53,54,55].
Another hypothesis is that the small bowel is likely a key site of amplification of the systemic inflammatory response, where blockage of ACE2 causes increased levels of angiotensinogen and hyperactivation of the renin–angiotensin system, leading to a shutdown of the amino acid transporter BA0T1 and a subsequent lack of cellular tryptophan, which leads to decreased secretion of antimicrobial peptides and gut dysbiosis [56].
Recent studies have observed that patients with COVID-19 develop a dysbiotic microbiota of the gut [24,57,58,59,60,61] (Figure 2). One study investigated changes in fecal microbiomes of patients with SARS-CoV-2 infection during hospitalization and associations with severity and fecal shedding of the virus [24]. Patients with COVID-19 had significant alterations in their fecal microbiomes, characterized by enrichment of opportunistic pathogens and depletion of beneficial commensals at the time of hospitalization and at all time points during hospitalization.
Depleted symbionts and gut dysbiosis persisted throughout hospitalization, even after clearance of SARS-CoV-2 and resolution of respiratory symptoms. Throughout hospitalization, Bacteroides dorei, Bacteroides taiotaomicron, Bacteroides massiliensis, and Bacteroides ovatus, which downregulate the expression of ACE2 in the murine gut, correlated inversely with SARS-CoV-2 load in fecal samples from patients. Interestingly, the baseline abundance of specific microbes in the gut, such as Coprobacillus, Clostridium ramosum, and Clostridium hathewayi, correlated with COVID-19 severity. There was an inverse correlation between the abundance of Faecalibacterium prausnitzii, an anti-inflammatory bacterium, and disease severity.
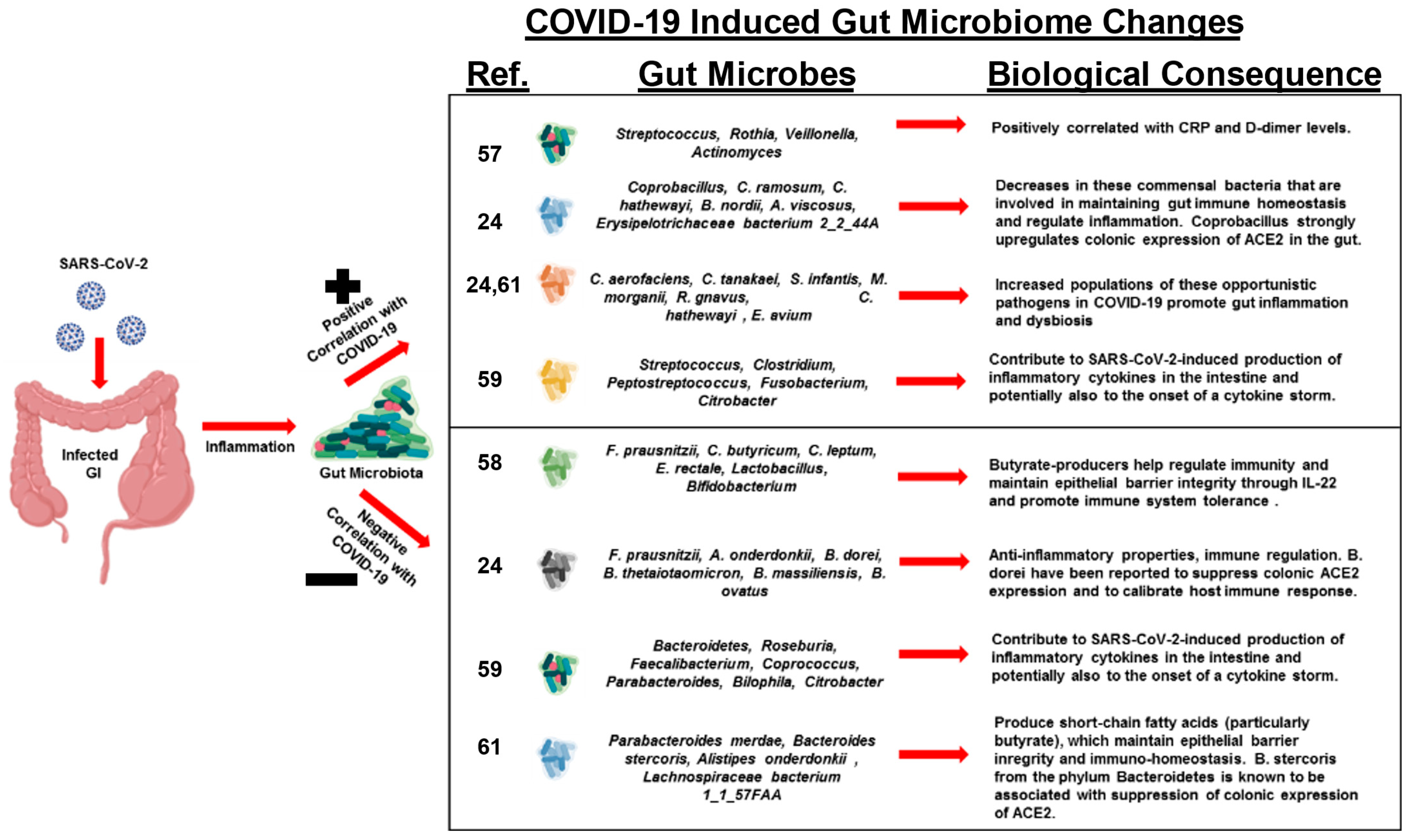
Another study investigated the changes in abundance of ten predominant intestinal bacterial groups in COVID-19 patients to establish a correlation between these bacterial groups and clinical indicators of pneumonia [58]. The results indicate that dysbiosis occurred in COVID-19 patients, and changes in the gut microbial community were associated with disease severity and hematological parameters. The number of common opportunistic pathogens, Enterococcus (Ec) and Enterobacteriaceae (E), were shown to be increased in COVID-19 patients, especially in the critically ill. Observations suggest that these bacterial groups can serve as diagnostic biomarkers for COVID-19 and that the Ec/E ratio can be used to predict death in critically ill patients.
The next study investigated the transcriptional activity of SARS-CoV-2 and its association with fecal microbiome alterations in COVID-19 patients [61]. Even in the absence of gastrointestinal (GI) symptoms, some patients continued to display an active viral infection signature up to 6 days after clearance of SARS-CoV-2 from respiratory samples. Fecal samples with a high SARS-CoV-2 signature had higher abundances of the bacterial species Collinsella aerofaciens, Collinsella tanakaei, Streptococcus infantis, and Morganella morganii. The gut microbiota of patients with active SARS-CoV-2 GI infection was characterized by enrichment of opportunistic pathogens, loss of salutary bacteria, increased functional capacity for nucleotide and amino acid biosynthesis, and carbohydrate metabolism. This study provides evidence for a dormant, prolonged GI infection by SARS-CoV-2, even in the absence of GI symptoms and after recovery from respiratory infection of SARS-CoV-2.
COVID-19 patients have also been shown to have significantly reduced bacterial diversity, a higher relative abundance of opportunistic pathogens, such as Streptococcus, Rothia, Veillonella, and Actinomyces, and a lower relative abundance of beneficial symbionts [57,59]. Levels of Fusicatenibacter, Romboutsia, Intestinibacter, Actinomyces, and Erysipelatoclostridium showed high accuracy for distinguishing COVID-19 patients, which suggests the potential value of the gut microbiota as a diagnostic biomarker and therapeutic target for COVID-19. Reduced bacterial diversity in the gut has been associated with acute and long-term metabolic effects and disease propensities.
In patients with COVID-19, the abundance of butyrate-producing bacteria, such as Faecalibacterium prausnitzii, Clostridium butyricum, Clostridium leptum, and Eubacterium rectale, decreased significantly, and this shift in the bacterial community may help discriminate critically ill patients from patients that have a more moderate disease state [58]. Fecal samples from patients with low-to-no SARS-CoV-2 infectivity had higher abundances of SCFA-producing bacteria, Parabacteroides merdae, Bacteroides stercoris, Alistipes onderdonkii, and Lachnospiraceae bacterium 1_1_57FAA [61].
Importantly, butyrate-producing bacteria are of critical importance in maintaining gut barrier integrality. SCFAs have been implicated in signaling for IL-22, which maintains gut and lung epithelial barrier integrity [62]. Butyrate and other SCFAs regulate inflammation by macrophages in the intestine and promote the Warburg effect, which metabolically constrains the neoplastic cells [63]. CRC has a metabolic dependence on anaerobic glycolysis; thus, fatty acid oxidation is limited. Given this metabolic state, butyrate begins to accumulate in the cytoplasm of cancerous colonocytes, allowing this SCFA to act as an HDAC inhibitor, sensitizing the cancerous colonocytes to apoptotic pathways [64,65]. Specifically, a reduction in butyrate-producing bacteria and gut dysbiosis contributed to less IL-22 production, which is integral to maintain both gut and lung epithelial barrier integrity [66] (Figure 3A,B).
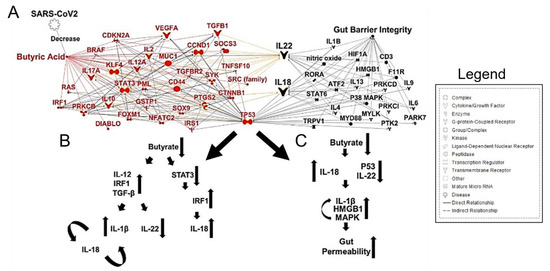
Fusobacterium nucleatum bacteremia has also been observed in severe COVID-19 cases and shown to colonize colon mucus with associated mucosal inflammation [67].
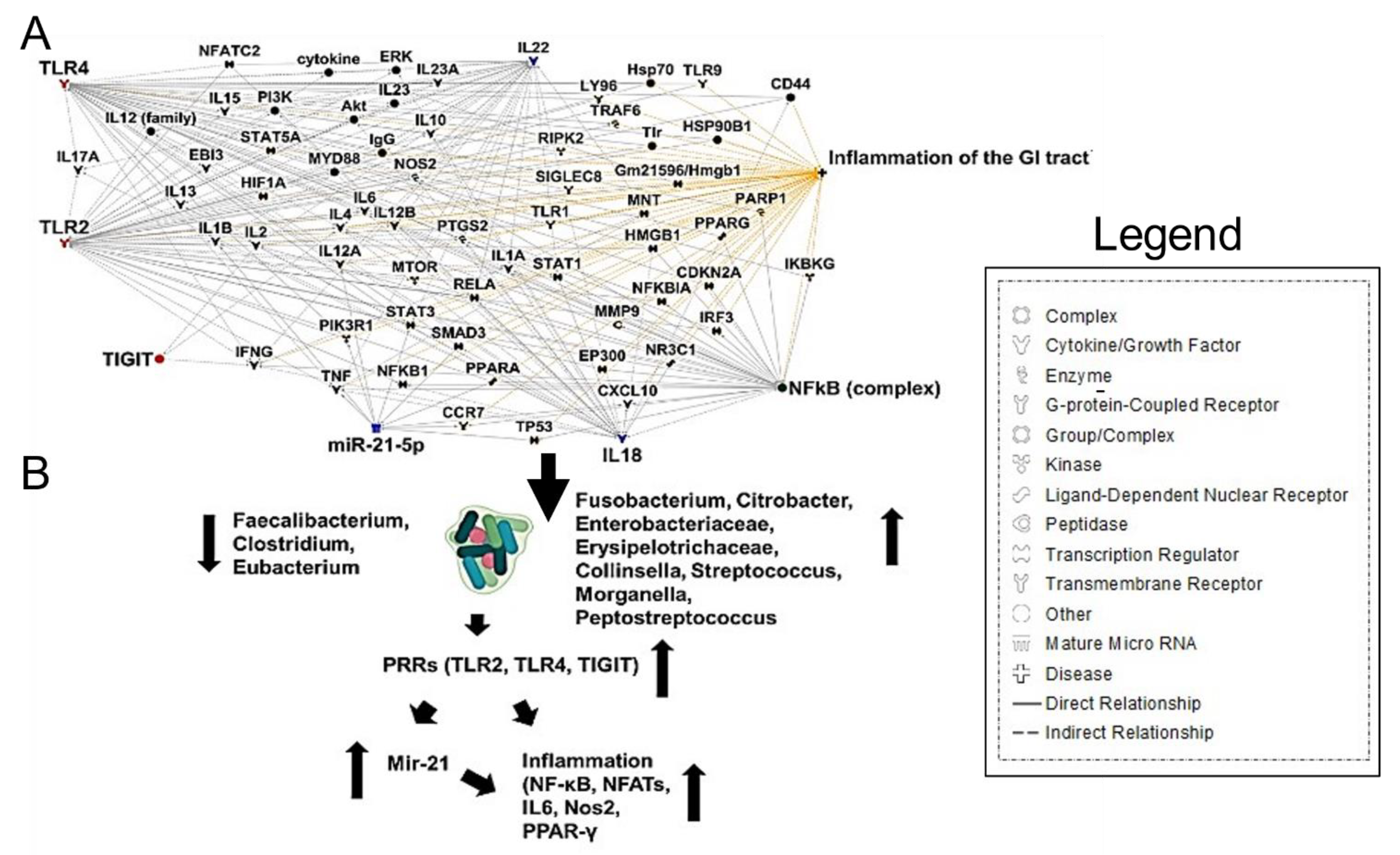
Along with the immune dysfunction seen in COVID-19, F. nucleatum causes immunosuppression by the action of one of its surface proteins, Fap2, which has been recognized as a galactose-binding lectin that can interact with the T cell ITIM domain (TIGIT) inhibitory receptor [68]. Furthermore, F. nucleatum was shown to interact with Toll-like receptors 2 and 4 signaling, resulting in increased microRNA-21 (miRNA21) expression, which promotes NF-kB induction, contributing to inflammation and cell proliferation [68] (Figure 4A,B). The intestinal inflammatory cytokine IL-18 was also shown to be elevated in the serum of COVID-19 patients [59]. These results indicate that gut microbiota composition changes contribute to the SARS-CoV-2-induced inflammatory cytokines production in the intestine.
Interestingly, one study also investigated changes in the fecal fungal microbiomes (mycobiome) of COVID-19 patients [69]. Patients with COVID-19 had significant alterations in their fecal mycobiomes, characterized by increased proportions of opportunistic fungal pathogens, Candida albicans, Candida auris, and Aspergillus flavus. Two respiratory-associated fungal pathogens, A. flavus and Aspergillus niger, were detected in fecal samples from a subset of patients with COVID-19, even after clearance of SARS-CoV-2 from nasopharyngeal samples and resolution of respiratory symptoms. These results are striking and re-enforce the commensal nature of all microbes, which may encompass bacteria, fungi, protists, and viruses alike.
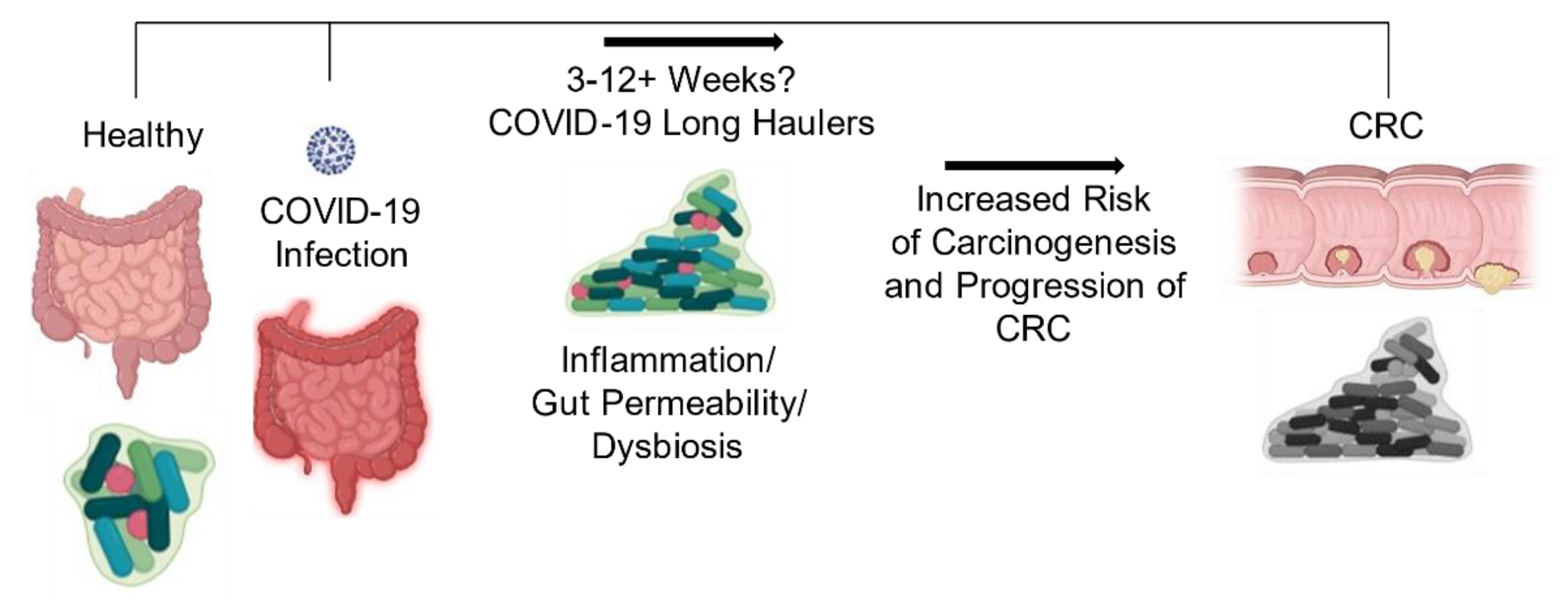
There are increasing reports of persistent and prolonged effects after COVID-19, now termed “long haulers”, a syndrome characterized by persistent symptoms and delayed or long-term complications beyond 4–6 weeks from the onset of symptoms [70,71]. This may be caused by the observed post-acute COVID-19 syndrome seen in some cases, where immune dysregulation and gastrointestinal symptoms are still ongoing even after clearance of SARS-CoV-2, thus impacting the status of the patient’s microbiome [72,73]. Due to the recent nature of the COVID-19 pandemic, the time of restoration of normal gut microbiota has yet to be thoroughly investigated, so the timeline and long-term effects of dysbiosis are unknown and require further studies.
A well-known respiratory infection phenomenon is gut-barrier dysfunction, which is correlated with a more severe clinical course of the disease [74]. Clinically, it has been shown that modulating the gut microbiota can slow early influenza virus replication in lung epithelia, resulting in reduced enteritis and ventilator-associated pneumonia [74]. Expansion of Bifidobacterium species upon and after influenza infection, for example, has been shown to be protective from the inflammation that could cause dysbiotic events [75,76]. Currently, there is no direct clinical evidence that the modulation of the gut microbiota plays a therapeutic role in the treatment of COVID-19. However, clinical trials of probiotics against COVID-19 are underway [77]. Probiotics can help patients maintain immune homeostasis in the gut and avoid overactivation of the immune response by decreasing proinflammatory signaling and maintaining gut barrier integrity.
The human immune system has evolved to cope with the presence of microorganisms both inside our body and out [78]. This fact means that most infectious diseases caused by viruses or bacteria are self-limiting [79,80]. This could be one possible reason for the asymptomatic or mild nature of most COVID-19 cases [81]. These microorganisms can serve as a source of metabolites, such as essential amino acids and fatty acids, for the human body [82]. In a state of gut dysbiosis, a transient nutritional supply excess may occur, as both the microorganisms and the damaged host tissue will be degraded and become a possible source of nutrition. This may promote hyper inflammation during acute infection [83] and possibly lead to chronic diseases, such as cancer [84]. For example, some of the excess nutrition from damaged tissue, together with the excess nutrition from a dysbiotic gut microbiome, could be turned into lipid intermediates, causing lipotoxicity and further tissue damage [85]. This highlights how the nutritional state can impact a person’s health and disease outcome. As such, food intake restriction could offer a promising way to control chronic inflammation [86].
Together, these collective observations warrant greater investigation into the relationship between changes in the gut microbiota that encompass all microorganisms due to COVID-19 infection and associations with increased risk for CRC development or progression (Figure 5). Given the widespread incidence of this virus and that CRC is the third most common cancer diagnosed in the United States in both men and women, studies investigating the associated effects of these two diseases are in great need. Upon elucidating answers to these questions, it may be possible to develop effective probiotic therapies that maintain a more homeostatic gut environment during the disease, which could help improve CRC survival rates.
reference link : https://www.mdpi.com/2072-6694/13/11/2676/htm



{kind=link}