









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


For the study, published April 1 in Molecular Psychiatry, senior author Dr. Dilek Colak, assistant professor of neuroscience at the Feil Family Brain and Mind Research Institute at Weill Cornell Medicine, and her colleagues grew astrocytes from the stem cells derived from patients with autism and transplanted them into healthy newborn mice.
They found that after the transplant, the mice developed repetitive behaviors, a hallmark symptom of autism spectrum disorders (ASD), but they did not develop the social deficits associated with the disease. The mice also developed memory deficits, which are commonly seen in ASD but are not a core characteristic of the disease.
“Our study suggests that astrocyte abnormalities might contribute to the onset and progression of autism spectrum disorders,” said Dr. Colak, who is also assistant professor of neuroscience in pediatrics and a member of the Drukier Institute for Children’s Health. “Astrocyte abnormalities may be responsible for repetitive behavior or memory deficits, but not other symptoms like difficulties with social interactions.”
Most studies of autism spectrum disorders have focused on the role of neurons, a type of brain cell that relays information in the brain. But other brain cells, called astrocytes, help regulate the behavior of neurons and the connections between them. Genetic mutations linked to autism spectrum disorders are likely to affect various types of cells in the brain differently, Dr. Colak said. Post-mortem studies had already revealed abnormalities in astrocytes in the brains of patients with autism spectrum disorders.
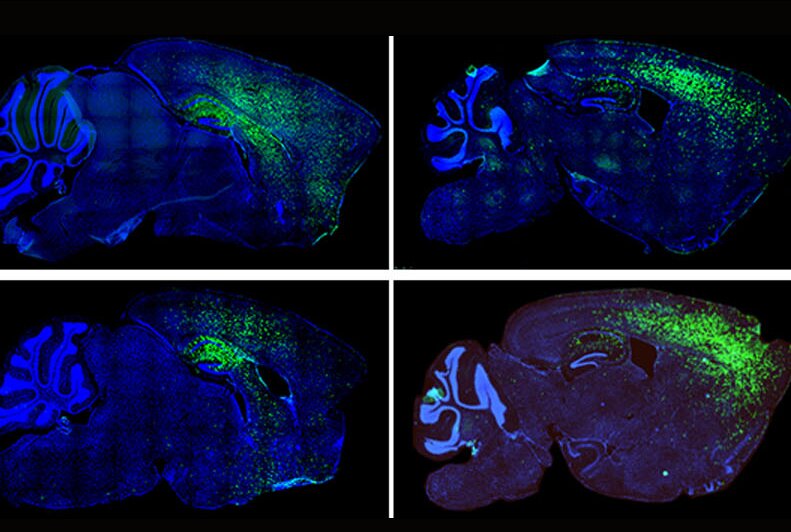
To determine if astrocytes might be involved early in the disease, the team obtained stem cells derived from patients with autism spectrum disorders, coaxed them into developing into astrocytes in the laboratory, and transplanted them into the brains of otherwise healthy newborn mice, creating a human-mouse chimera.
Using a microscopic technique called two-photon imaging, they observed excessive calcium signaling in the transplanted human astrocytes in the brains of mice, explained co-lead author Dr. Ben Huang, instructor of neuroscience in psychiatry at Weill Cornell Medicine.
To determine if the increased calcium signaling was causing the mice’s behavioral symptoms, the team infected astrocytes grown from ASD patient stem cells in the laboratory with a virus carrying a fragment of RNA designed to reduce calcium signaling to normal levels. When they transplanted these astrocytes into the mice, the animals did not develop memory problems.
“Future therapies for autism might exploit this finding by using genetic tools to limit extreme calcium fluctuations inside astrocytes,” said co-lead author Megan Allen, a postdoctoral associate in neuroscience in the Feil Family Brain and Mind Research Institute at Weill Cornell Medicine.
“It is important to determine the roles of specific types of brain cells, including astrocytes, in neurodevelopmental and neuropsychiatric diseases,” she said.
Glia: From Passive Glue to Excitable Cells
When neuroglia were first described, there was considerable debate as to whether neuroglia were a connective tissue or a true population of cells (Somjen, 1988). While this issue was resolved in the late 1800s, little attention was paid to the role of glia in neurophysiology for nearly a century. During this period, neuroscientists generally considered glia as chemical and physical insulators that enabled neurons to carry out the diverse functions of the brain.
This view was reinforced by the findings of early neurophysiologists who impaled glial cells with sharp electrodes and found only passive membrane currents. Given the large fraction of brain contributed by glia, the prevailing view until the early 1970s was that over half of the mammalian brain was, in effect, silent.
This view began to change as investigators found that glial cells in culture exhibited a large number of G protein-coupled receptors (GPCRs) linked to a diverse array of intracellular signaling cascades (McCarthy and de Vellis, 1978, van Calker and Hamprecht, 1981, van Calker et al., 1978).
Concerns about the expression of GPCRs by glia being a “culture” phenomenon were put to rest as it was demonstrated that glia in situ and in vivo also express GPCRs (Porter and McCarthy, 1997). Today, it is generally accepted that glia throughout the brain and spinal cord as well as peripheral glia residing within ganglia and aligning axons express members of most of the different families of GPCRs known to be expressed by neurons (Porter and McCarthy, 1997).
Stimulation of these GPCRs evokes a variety of glial cell responses, the most studied of which is elevation of intracellular calcium (Ca2+) concentration that is widely considered a form of glial excitability. The question is no longer whether glia exhibit GPCRs, but under what conditions are these GPCRs activated and what is the role of glial GPCR-mediated signaling in neurophysiology?
Astrocytes Are the Predominant Glial Cell Type in the Central Nervous System
Like the term “neuron,” glia refers to a diverse set of cell types that are likely to carry out distinct functions in neurophysiology. There are four major groups of glial cells in the nervous system:
(1) Schwann cells and oligodendrocytes, which produce and wrap layers of myelin around axons in the peripheral and central nervous systems, respectively;
(2) microglia, the immune cell type of the nervous system, which participate in inflammatory responses;
(3) nerve/glial antigen 2 (NG2)-positive glia, which include oligodendrocyte and astrocyte progenitor cells as well as NG2+ cells that persist in the mature brain; and
(4) astrocytes. Astrocytes are found throughout the brain and spinal cord and, on the basis of number, surface area, and volume, are the predominant glial cell type.
There are many distinct subsets of astrocytes that can be distinguished on the basis of their morphology and biochemical characteristics. For example, Mueller glia in the retina and Bergmann glia in the cerebellum are generally grouped with astrocytes because of their expression of glial fibrillary acidic protein (GFAP) but exhibit striking differences in morphology, pharmacology, and physiology (Grosche et al., 1999, Grosche et al., 2002, Metea and Newman, 2006, Pinto and Gotz, 2007).
It is likely that, even within a localized brain region, adjacent astrocytes that appear identical morphologically and immunocytochemically may vary in their expression of GPCRs and their response to activation of GPCRs. While such diversity is generally accepted when considering neurons, it is rarely taken into account when interpreting data derived from astrocytes.
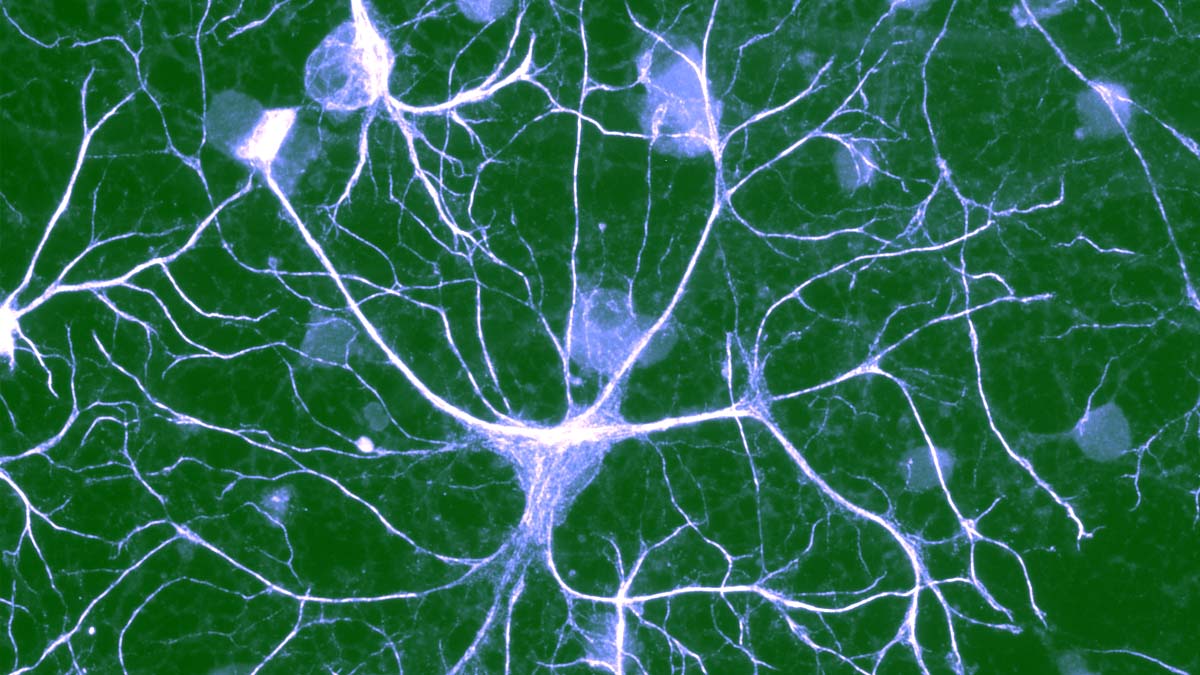
Protoplasmic astrocytes are the most common type of astrocytes. These cells exhibit a very complex morphology and contact most, if not all, other cell types in the brain and spinal cord. The morphology of an astrocyte resembles a bush with processes radiating out from a central cell body (Figure 1A).
Within the CA1 stratum radiatum of the hippocampus, an individual astrocyte has a soma diameter of 7–9 μm and, with its fine processes, occupies a volume of ∼66,000 μm3 (Bushong et al., 2002). Interestingly, individual astrocytes tend to occupy distinct, nonoverlapping domains (Figure 1A) (Bushong et al., 2002).
The fine processes of an individual astrocyte are connected to one another through reflexive gap junctions and to other astrocytes via gap junctions at their boundaries. Patch-clamping a single astrocyte with an electrode filled with a gap-junction-permeable dye rapidly leads to the filling of hundreds, if not thousands, of astrocytes (Konietzko and Muller, 1994).
Astrocytes likely function as a syncytium contacting essentially all other cellular elements in brain, including neurons, oligodendrocytes, NG2+ cells, microglia, and vasculature. A striking feature of astrocytes is that processes from a single astrocyte can envelop approximately 140,000 synapses (Figure 1B) (Bushong et al., 2002), while >99% of the cerebrovascular surface is ensheathed by astrocyte processes (Kacem et al., 1998, Rama Rao et al., 2003, Simard et al., 2003, Haydon and Carmignoto, 2006, Takano et al., 2006).
In addition to the diversity among astrocytes, there may be substantial diversity within individual astrocytes with respect to interactions with the local environment. For instance, it is possible that, within a single astrocyte, a subset of processes interacts autonomously with a neighborhood of neuronal synapses, while other regions of that astrocyte interact with different groups of synapses or with other cellular elements, such as the cerebrovasculature.
Further, under physiological conditions, these local regions of interaction (microdomains) of an astrocyte may not always communicate with one another. Understanding how the different microdomains of astrocytes interact with neighboring cellular elements will be critical to determining their role in neurophysiology and neuropathology.

reference link : https://www.cell.com/fulltext/S0896-6273(08)00751-4
Different Models of ASD
The most widely used and validated environmental animal model of ASD is the exposition to valproic acid (VPA). The VPA model is based on epidemiological studies which showed a high incidence of autism in a newborn child following exposure of the pregnant mother to this antiepileptic drug [14]. Furthermore, exposition to VPA results in numerous deficits in social behaviour and communication in rodents [15,16]. These behavioural alterations manifest from the early stages of development [17,18,19].
VPA is a short-chain fatty acid known for its anticonvulsive activity. It has been widely used for epilepsy as well as migraine and bipolar disorder treatment for several decades [20]. VPA has the ability to modulate neurotransmission and regulate GABA transmission and gene expression by inhibition of histone deacetylase activity [21].
Unfortunately, it turned out that VPA is also a potent teratogen leading to vast brain malformations in offspring when administrated during pregnancy. These abnormalities of normal neurodevelopment, in turn, result in developmental delay and autism [22]. Behavioural deficits induced by VPA exposure in humans are also consistently exhibited in rodents.
Another recognised risk factor of the occurrence of psychiatric disorders, besides VPA exposition or genetic mutations, is infection during prenatal life. Activation of the maternal immune system (MIA) caused by viruses or bacteria might lead to neuropsychiatric disorders in the offspring.
The MIA model can be divided into three phases. First, viruses or other pathogens activate the maternal immune system and lead to increased levels of proinflammatory cytokines. Consequently, the disturbed integrity of the placental barrier allows for the entrance of maternally derived proinflammatory factors into the fetal circulation. That evokes an inflammatory response in the developing brain of the fetus. Disturbed neurogenesis and central nervous system development will be the cause of neuropsychiatric disorders in the offspring [23].
Due to the nature and specificity of behavioural impairments and structural changes of the brain caused by inflammatory processes, inflammation-based animal models have been widely used to explore certain aspects of human disorders such as autism but also schizophrenia. Moreover, there are plenty of studies and meta-analyses, which have shown that maternal infection increases the risk of the occurrence of neuropsychiatric disorders in children [24].
To study the mechanism behind the association of MIA and ASD occurrence in offspring, pregnant rodents have to be infected with: the lipopolysaccharide (LPS), which leads to a bacteria-like infection; poly (I:C), double-stranded RNA analogue, causing virus-like infection; or influenza virus. However, the chance of developing autism is greater after viral infection, especially when it happens in the first trimester of pregnancy [25]. Poly (I:C) injection to pregnant rodent females is considered to have a stronger construct validity than LPS [23]. Rodents prenatally exposed to poly (I:C) treatment have several socio-communicative features of autism, such as decreased number of social play episodes and deficits in communication, abnormal vocalisation, as well as altered cytokines levels or neuronal density within the brain [13].
About 15% of ASD cases have a genetic component such as coding sequence mutations, chromosomal rearrangements, or copy number variants [26]. For example, Fragile X syndrome is a genetic condition that results in a range of cognitive impairments and is the most common monogenic cause of autism in human patients and animal models [27]. Actually, Fragile Mental Retardation 1 (Fmr1) knock-out (KO) is one of the best preclinical genetic models associated with autistic traits.
Altered behaviour in clinical ASD cases and preclinical models is associated with a plethora of functional and morphological changes in the brain. Impairment of synaptic connectivity is a well-known cause of ASD. Fmr1-KO mice exhibit disturbed organisation and morphology of dendrites and synapses, as well as impaired structure of the cortex, cerebellum, and hippocampus, which is known to be a manifestation of improper neuron–glia connectivity.
In addition to the variety of mutations associated with ASD, disturbances of histone 3 lysine 4 (H3K4) methylation during prenatal development may also be the cause of neurodevelopmental disorders, including autism. Lysine-specific demethylase 5C—KDM5C is able to remove methyl groups from tri- and dimethyl H3K4, which have an impact on proper dendritic growth and survival, and development of neurons [28].
Mutation of KDM5C is present in children with Fragile X syndrome. Furthermore, it has been shown that iKdm5C-KO animals have impaired social behaviour, cognitive abnormalities, memory deficits, and enhanced aggressive behaviour [29]. However, recent studies have shown that using some FDA approved drugs such as histone deacetylase inhibitor suberanilohydroxamic acid (SAHA) may force the KDM5C expression and lead to improvement of neuronal maturation and abnormal behaviour [30]. Further studies focused on using SAHA may be very helpful in the therapy of ASD.
In 2017 Pavăl [31] presented the dopamine (DA) theory of autism. He postulated that DA signalling might be involved in the symptomology of ASD. A lot of studies have shown changes in levels of DA and changes of phasic striatal DA events in different brain structures in ASD patients. Moreover, antagonists of dopamine receptors diminished some symptoms of autism (reviewed in [32]). It has been also demonstrated that stimulation of the nigrostriatal pathway using optogenetic tools influences social and repetitive behaviour in mice [33].
Furthermore, mutations of genes encoding dopamine transporters (DAT T356M) lead to hyperlocomotion, reduced social approach and deregulated DA metabolism and synthesis [34] in rodents. García-Domínguez and colleagues [35], in the most recent study, have shown that selective deletion of Caspase-3 gene in the dopaminergic system leads to spatial changes of DA levels, and most importantly, Casp3-deficient mice exhibited impaired social interaction, restrictive interests and repetitive behaviours—the core symptoms of autism. It is worth mentioning that spatio-temporal changes in the dopaminergic system associated with ASD are dynamic; therefore, further research is needed to solve the possible role of DA in ASD.
The models described here are not perfect, but each of them provides valuable information and data needed to understand the mechanism involved in autism. A short list of the chosen animal models of autism is provided in Table 1.
Table 1
A short list of the chosen animal models of autism.
| Models | Basis | Mechanism | Observed Behavioral Alterations | Bibliography |
|---|---|---|---|---|
| LPS | inflammation | increased levels of proinflammatory cytokines in the brain caused by bacterial infection | disturbed communication, altered social behaviour, increased repetitive behaviour | [36] |
| POLY (I:C) | inflammation | increased levels of proinflammatory cytokines in the brain caused by viral-like infection | disturbed communication, altered social behaviour, increased repetitive behaviour | [19,37] |
| VPA | chemical pharmacological | a potent teratogen leading to vast brain malformations, inhibition of histone deacetylase, GABAergic signalling disruption | disturbed communication, altered social behaviour, increased repetitive behaviour, anxiety | [16,38] |
| PPA | inflammation | dietary and gastrointestinal agent initiating neuroinflammation and gliosis | altered social behaviour and cognition | [39] |
| FMR1 | genetic | mGluR5 hyperactivation | disturbed communication, altered social behaviour, seizures, cognitive impairments | [40] |
| SHANK1 | genetic | disruption in synaptic transmission | disturbed motor function, memory, altered communication, no changes in social interactions | [41,42] |
| SHANK3 | genetic | disruption in synaptic transmission | disturbed communication, altered social behaviour, altered learning and memory | [43] |
| TSC1, TSC2 | genetic | altered mTOR signalling | altered communication, altered memory and learning | [44] |
| NLGN1,2,3,4 | genetic | disturbed synaptogenesis | increased stereotypic behaviour, altered sociability | [45] |
| RELN | genetic | disturbed neuronal positioning and synaptogenesis | disturbed communication, altered social behaviour | [46] |
| CNTNAP2 | genetic | decreased glutamate receptor expression and transmission | disturbed communication, altered social behaviour, hyperactivity | [47] |
| CACNA1C | genetic | disruption in synaptic transmission | disturbed communication, altered social behaviour | [48] |
| GABRB3 | genetic | altered GABAergic signalling | disturbed communication, altered social behaviour, hyperactivity, altered cognition | [49] |
| UBE3A | genetic | disruption in synaptic transmission | disturbed communication, altered social behaviour, seizures, altered cognition | [50] |
Astrocytes as Important Players in the CNS
Astrocytes are the main cells responsible for maintaining brain homeostasis. They are also involved in mechanisms associated with the metabolism and signalling between other cells in the central nervous system such as neurons, oligodendrocytes and microglia. To maintain these functions, astrocytes express a lot of different molecules and receptors. Moreover, astroglia have a plethora of different transporters and pumps, which are essential for proper signalling between the brain cells.
In opposition to neurons, astrocytes are not electrically excitable, so they “talk” using the fluctuation of different ions such as calcium, sodium and potassium. This intracellular transport of the ions is possible thanks to the integration of the astrocytes into the syncytia in which the astroglial membranes are connected by the gap junctions (small channels formed by the connexin (Cx) protein family, particularly Cx30 and Cx43).
Ion homeostasis is a critical function of astroglia. Astrocytes regulate potassium ions and water flow in the extracellular space and between neuronal cells using inward rectifying K+ channels (Kir) for buffering K+ and water channels—Aquaporin4 (Aqp4)—needed for clearing the cellular debris by the glymphatic system [74]. Altered homeostasis of astrocytic water channels and K+ ions in the brain lead to altered balance between neuronal excitation and inhibition. Even small alterations in extracellular K+ concentration may produce hyperexcitability of neurons.
Reduced Kir4.1 expression (but not other K+ channels) increases extracellular K+. Additionally, conditional knock-out of Kir4.1 depolarises glial membranes and inhibits potassium and glutamate uptake [75]. Furthermore, it has been shown that loss of water channels such asAqp4 present on astrocytes may lead to an impaired K+ buffering and consequently to increased excitation of neurons which underlies many neuropsychiatric disorders, including autism. The scheme depicting alterations in astroglia–neuron interactions (along with their probable causes) that possibly contribute to the development of autism is shown in Figure 2.

The possible role of astrocytes in the development of autism. Genetic mutations, prenatal exposition on some drugs and prenatal inflammation affect astrocytic morphology and functions. It may result in alterations of synapse functions, imbalanced homeostasis between excitation and inhibition and dysregulation of the nervous system. This, in turn, may lead to autistic-like behavioural changes. D-Ser: D-serine; GLAST, GLT-1: glutamate transporters; Aqp4: aquaporine 4; Kir4.1: inward rectifying K+ channels.
It has been known for a long time that astrocytes are essential for maintaining proper brain activity. Blocking astrocytic metabolism by fluorocitrate leads to seizures and may be the cause of obsessive/compulsive behaviour in rodents [76]. Astrocytes are responsible for the clearance and transport of glutamate, which is possible due to the presence of glutamate transport proteins GLAST and GLT-1 on astrocytic membranes. Abnormalities of glial cells regarding glutamate metabolism may lead to behavioural impairments in animals.
Aida and colleagues (2015) [77] found that GLT-1 deficiency increases neuronal excitation and leads to excessive repeated behaviour, including self-injuring in mice. That should not be very surprising knowing that GLT-1 accounts for more than 90% of glutamate uptake in the brain. These findings were recently confirmed by another research group [78,79].
Furthermore, postnatal disturbance of glia proliferation may lead to hyperactive behaviour in open field tests and alterations of social interactions in rats [80]. In addition to the abovementioned glutamate transporters, the amino acid neurotransmitter cycle is also supported by glutamine synthetase (GS). GS converts glutamate into glutamine which will then be used by neurons.
As it turns out, GABA-ergic neurons are more dependent on astrocyte glutamine and thus more sensitive to decreased GS activity than excitatory neurons, so lack of the astrocytic GS may lead to the altered functioning of the inhibitory neurons. Furthermore, inhibiting the astrocytic GABAB-Gi pathway in the striatum leads to the enhanced attention in mice and reduces behavioural hyperactivity in mice [81].
There is still growing evidence that not only neurons but also astrocytes are involved in memory formation. This is possible due to the release of the astrocytic transmitters such as D-serine and ATP following the activation of astroglia [82]. Astroglia may contribute to cognition processes not only by modulating synapses but also by activating the neighbored neurons by supplying them with lactate. Since the last decade, the pivotal role of astrocytic lactate in memory formation and learning has consequently been confirmed. Moreover, it has been shown that the lack of lactate transportation may even result in amnesia in rodents (reviewed in [83]).
While there are indications that astrocytes underlie the mechanisms leading to psychiatric disorders such as autism, and their altered functioning may lead to behavioural changes in rodents, there is still little research on this topic, especially regarding social behaviour and communication, which should be the next step in understanding the mystery of cooperation between neurons and astrocytes.
Shape Changes of Astroglia
Astrocytic processes tend to change their morphology in response to CNS inflammation, epilepsy, injuries, and neurodegenerative disease as well as during the normal, non-pathological interactions with neurons. These morphological changes of astroglial silhouettes may act as excellent indicators of the current state of the brain; a detailed morphological analysis is able to show changes already in their initial stages. Most astroglia is generated by the 16th postnatal day [84].
However, through this period of time, they tend to change their morphology from immature, characterised by a simpler silhouette with quite short, uncomplicated processes, to mature, characterised by a fine, largely ramified profile. At postnatal day 14, all astrocytes should be characterised by almost-mature morphology, even though they do not reach full maturity until postnatal day 30 [85]. This process of astrocytic maturation is temporally correlated with the maturation of the synapses and the expression of some glutamate receptor subunits [86], as well as their identity may be overwritten by surrounding neurons [87].
During synaptogenesis, astrocytes’ morphology becomes more complex, with small but very important perisynaptic processes which are engaged in establishing “tripartite synapses” with neurons. Astrocytic perisynaptic processes are very rich in a plethora of receptors, adhesion molecules and ion channels which are crucial for maintaining functions of synapses (reviewed in [88]).
Astrocytes, based on their morphology, can be divided into two main sub-types, namely, protoplasmatic and fibrous. Fibrous astrocytes are distributed in white matter and have an elongated shape with numerous, long processes. Protoplasmatic astrocytes reside in grey matter and they are often called bushy because of their shape and highly ramified processes.
The processes are sandwiched between neurons for the purpose of being in close contact with synapses. Recent studies have shown that protoplasmatic astrocytes are diverse and manifest layer-specific molecular and morphological differences. Lanjakorsiripan et al. (2018) [89] demonstrated that astrocytes in mice cortex have different shapes in different cortical layers. For example, astrocytes from layer II/III tend to elongate radially in contrast to astroglia in layer VI, which elongate tangentially. Moreover, using 3D Sholl analysis, they found that astrocytes in layer II/III have more intersections than layer VI astrocytes.
The shape of astrocytic silhouette turned out to be associated with their function in the cortex. Although most of the genes expressed on astrocytes are similar, some of these genes, mainly engaged in synaptogenesis and metabolism, were enriched in astroglia in the upper cortical layers (Dio2, Mertk, Slc1a3). Genes highly expressed in deeper layers were mostly associated with proliferation (Cxcr7) or astrocyte–neuron communication (Gfap).
Furthermore, the Lanjakorsiripan group also found that the formation of layer-specific characteristics of cortical astrocytes is strongly dependant on neurons. Without neuronal signalling astrocytes failed to form the laminar organisation. There is a strong connection between astroglia and neurons, and they should not be studied separately but rather together as one coexisting/symbiotic net.
Unfortunately, there is little precise research on the shape of astrocytes in ASD. This is quite surprising because there are available tools to perform the detailed analysis of cell morphology. Using ImageJ software [90], different research groups were able to distinguish between different types of glial cells, including all intermediate states of the cell [91].
Parameters which can be used to quantify morphological changes of glia can be associated with a number of processes (sum of intersections, maximal number of intersections, ramification index (obtained with Sholl analysis)). It is also possible to describe the size and shape of the cell (convexity, solidity, etc.).
Moreover, using fractal dimension and lacunarity we are able to study the complexity of the cell. Since astrocytes are very heterogeneous plastic cells and they can change their morphological and functional properties throughout their lifetime, it seems important to have a closer look at the morphological changes in astrocytes in further research. Nevertheless, it should be taken into account that the change in the shape of astrocytes and the gain or loss of some protective or harmful functions may occur simultaneously. The impact of these changes on the course of various processes/diseases will change depending on the balance between the protective and harmful alterations that have occurred [92].
reference link : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8583956/
More information: Megan Allen et al, Astrocytes derived from ASD individuals alter behavior and destabilize neuronal activity through aberrant Ca2+ signaling, Molecular Psychiatry (2022). DOI: 10.1038/s41380-022-01486-x



{kind=link}