









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Some of the most fundamental questions in evolution remain unanswered, such as when and how extremely diverse groups of animals – for example reptiles – first evolved.
For seventy-five years, adaptive radiations – the relatively fast evolution of many species from a single common ancestor – have been considered as the major cause of biological diversity, including the origins of major body plans (structural and developmental characteristics that identify a group of animals) and new lineages.
However, past research examining these rapid rates of evolution was largely constrained by the methods used and the amount of data available.
In a paper out today in Nature Communications, a research team lead by Harvard University examined the largest available data set of living and extinct major reptile groups (such as marine reptiles, turtles, lizards, and the ancestors of dinosaurs and crocodiles) to tackle the longstanding question of how adaptive radiations have shaped reptile evolution.
Using DNA information from modern species and hundreds of anatomical features from both modern and fossil species for statistical analysis, the study detected that periods of fast anatomical change during the origin of reptile groups often predate when those groups diversified into hundreds or thousands of species. This contradicts long-held ideas of adaptive radiation in evolution biology.

“Our findings suggest that the origin of the major reptile groups, both living and extinct, was marked by very fast rates of anatomical change, but that high rates of evolution do not necessarily align with taxonomic diversification” said first author Dr. Tiago Simões, Postdoctoral Fellow in in the lab of Stephanie Pierce, AssociateProfessor in the Department of Organismic and Evolutionary Biology at Harvard University.
Simões and Pierce revealed that rates of evolution and morphological variety in reptiles prior to the Permian-Triassic Mass Extinction – the biggest mass extinction of all time – were equally high, or even higher, than after the event.
As reptile species diversity was much lower during the Permian compared to Triassic, these results indicate that fast rates of evolution do not need to coincide with rapid taxonomic diversification as predicted by the classical theory of adaptive radiation. The two can be decoupled.
The team, which also included Ph.D. student Oksana Vernygora and Professor Michael Caldwell at the University of Alberta, further discovered that accelerated rates of evolution correspond to the origin of unique reptile body plans, but that very similar functional adaptations in reptiles can arise through varying rates of evolution.
“Surprisingly,” Pierce said, “reptiles that evolved similar protective armour like turtles or serpentine bodies like snakes, show radically different rates of evolution, indicating the origin and evolution of unique body plans is heterogeneous through evolution.”
“Our results also show that the origin of snakes is characterized by the fastest rates of anatomical change in the history of reptile evolution,” said Simões. “But, that this does not coincide with increases in taxonomic diversity [as predicted by adaptive radiations] or high rates of molecular evolution.”
The mismatch between morphological and molecular evolution supports the idea that protein coding DNA sequences do not seem to be correlated with broad-scale changes in anatomy.
Although much more research is needed to understand how body plans evolve, the team hypothesizes that non-protein coding regions of the genome may be responsible for rapid morphological change, as these parts are more free to mutate and take on new functional roles.
“It is clear to us that to advance our understanding of the major patterns in evolution we need further studies capable of measuring phenotypic and molecular evolutionary rates, times of origin, and phenotypic diversity across large timescales” said Simões.
Simões and colleagues continue to develop new methods and are expanding their data set back in time to look at the origins of amniotes, the group that includes both reptilesand mammals.
Of particular interest is pinpointing when in geological time these two groups of animals diverged and how extinction, diversification, and adaptation have shaped their evolutionary history over the last 300+ million years.
“I’m excited to continue my research to unravel the early evolutionary dynamics of the two most successful groups of animals on the planet,” Simões said. “I’m also focusing on improving available protocols to analyze morphological data and construct more robust evolutionary trees, including the timing of origin of major vertebrate lineages.”
Evolution of the reptiles
The reptiles make up a huge group of fossil and living vertebrates, ranging in size from tiny thread snakes to sauropod dinosaurs, which are the largest animals ever to have lived on land.
Through time reptiles have evolved into unique forms, such as turtles, snakes, and dinosaurs, but they also have taken on the appearance and habits of other vertebrate groups, such as sharks and dolphins. As with other animal classes, reptile groups that are thought to share a common ancestor are known as clades.
The application of cladistics has changed ideas about how organisms should be classified. For instance, because they are thought to be descended from small bipedal dinosaurs, birds are now included with the reptiles. But synapsids (once called mammal-like reptiles) are classified with the mammals.
Because so many diverse animals are included under the term reptiles, they are difficult to define as a single group. Reptiles are amniotes, that is, they are tetrapods (four-legged vertebrates) with an amnion that surrounds and protects the developing embryo.
Reptiles other than birds and their immediate ancestors lack true feathers, and all of them lack true hair. Common (though varying) characteristics among reptiles include the fact that they cannot regulate their temperature
internally (with the possible exceptions of some dinosaurs and all birds), that they have an extensive covering of scales or bony plates or both (individual exceptions occur in many major groups), that they have a three-chambered heart (with the exceptions of crocodilians and, possibly, other archosaurs), and that they have 12 pairs of cranial nerves.
The major reptile groups considered here are Anapsida (“stem reptiles,” turtles, and other primitive groups), Euryapsida (the marine nothosaurs, plesiosaurs, placodonts, and ichthyosaurs), and Diapsida.
The last group includes the Lepidosauria (sphenodontians, such as tuatara and its fossil relatives; lizards; and snakes) and the Archosauria (pseudosuchians; crocodilians; pterosaurs, also known as “flying reptiles”; and dinosaurs).
Each of the three major reptile groups are defined on the basis of the number and position of large openings in the temporal region of the skull behind the eyes (relative to other skull bones).
Anapsid reptiles have no large openings in the temporal region of the skull and were the first stem to branch off the reptilian lineage. Euryapsid reptiles have a single temporal opening in the upper part of the skull. Diapsid reptiles have two large temporal openings, one above and one below a horizontal bony bridge.
Anapsida
The earliest reptiles are known from the early Pennsylvanian (323–317 million years ago, or mya). They were quite small and lizardlike in appearance, and their skulls, jaws, and tooth structures strongly indicate that they were insectivorous.
In fact, it is thought that they evolved in tandem with insect groups that were beginning to colonize the land. Some Pennsylvanian (323–290 mya) amphibians of the microsaur group also evolved into insectivores that were so superficially similar to early reptiles that, for a time, they were classified as such.
The amniote egg evolved in the earliest reptiles. This allowed for the first true occupation of the land by tetrapods, for the amniote egg allowed the embryo to develop in an aquatic microcosm until it was ready for terrestrial life; this paved the way for the huge adaptive radiation that eventually took place among the reptiles.
Robert Carroll of the Red-path Museum in Montreal, Canada, has pointed out that the earliest reptiles probably occupied the land before the amniote egg was developed fully. An analogy may be found in a few modern salamanders (small, somewhat lizardlike amphibians) that lay tiny non-amniote eggs in moist terrestrial places, such as under logs or in piles of damp leaves.
These eggs hatch into tiny replicas of the adults rather than going through a larval stage, such as occurs in frogs and other salamanders. The evolution of the amniote egg took place when the membranes within the eggs of the earliest reptiles became rearranged in the form of various sacs and linings and the outer membrane incorporated calcium into its structure to form a shell.
The calcareous (limey) shell afforded protection for the developing embryo and was porous enough to allow for the entrance and exit of essential elements, such as oxygen and carbon dioxide.
Food for the developing embryo was supplied in the form of an extra-embryonic sac full of yolk and a system of blood vessels that allowed the yolk to be transferred to the embryo.
The amnion itself formed a sac that contained a fluid within which the embryo was suspended. This provided a “private pond” (a term used by the late Harvard University scientist Alfred S. Romer, the world leader in vertebrate paleontology from the late 1940s until his death in 1973) for its occupant and kept the fragile embryonic parts from sticking together.
A unit composed of a part of the allantois and the membranous chorion next to the shell allowed for the absorption of oxygen and the excretion of carbon dioxide. A sac formed by the allantois stored the nitrogenous wastes excreted by the embryo. The origin of the amniote egg was one of the most important evolutionary events that ever occurred.
Turning to other anapsid reptiles, turtles were one of the first reptiles to branch off the amniote stem. Turtles are first known from the late Triassic (227–206 mya) but probably evolved in the Permian (290–248 mya).
Aside from protection, the turtle shell (which is essentially a portion of its skeleton turned inside out) has other critical functions. A large percentage of the red blood cells of most land vertebrates are formed in the marrow of the long bones.
Turtles, however, need sturdy legs to support their shells; thus, their limb bones are very dense, with little space, if any, for red blood cell-producing marrow.
It has been shown that the turtle shell is filled with canals and cavities where red blood cells are produced in quantity. Moreover, rather than being just an inert shield, the turtle shell is the site of calcium metabolism and is important in the process of temperature regulation, by absorbing heat during the basking (sunning) process.
Other attributes of turtles include the ability of some of them to absorb oxygen in the water through patches of thin skin on the body, in the lining of the mouth, or within the cloaca, a terminal extension of the gut wall.
Some turtles can freeze solid in the winter and thaw out in the spring with no harmful effects—a process that also takes place in various frog species. The ability of turtles to survive severe injuries is well known, and many species can exist in the absence of oxygen for long periods of time.
Leatherback turtles have a current-countercurrent blood flow similar to that of deep-sea mammals and can dive in the sea at great depths and remain active
in very cold temperatures. The incubation temperature of the eggs of many species of turtle determines the sex of the hatch-lings. In some cases “cold nests” produce females and “warm nests” produce males, but the opposite can also occur. Turtle eggs have large amounts of yolk compared with those of many other vertebrates. This “egg food” sustains the young during long incubation periods.
The origin of turtles is somewhat in doubt. An early anapsid with expanded ribs, Eunotosaurus, once was proposed as the ancestral form, but the skull of Eunotosaurus was not turtle-like. On the other hand, the body skeleton of early reptiles called procolophonids had a shell, and the skull was somewhat turtlelike.
Owenetta, a procolophonid found in the Upper Permian (256–248 mya) of South Africa has nine advanced characters in the skull and one in the humerus that are shared with Proganochelys, an unquestionable Triassic (248–206 mya) turtle. Owenetta lacks a shell, however; thus it has been suggested that the skull changes in turtle ancestors preceded those that led to the origin of the shell.
True turtles (order Testudines) are composed of three major suborders: the proganochelydians, the pleurodirans, and the cryptodirans.
The proganochelydians are the most primitive and are known from the late Triassic to the early Jurassic (206–180 mya). The shell is similar to that of modern turtles, except that it has extra bones and the head and limbs cannot be retracted effectively into it. The skull lacks teeth except for a few on the palate. The pleurodirans and cryptodirans have no teeth in the skull and can retract the head, neck, and tail into the shell.
In the pleurodires the neck swings sideways when it is retracted, so that the turtle looks out with only one eye. In the cryptodires the neck folds over itself when it is retracted, so that the turtle gazes with both eyes. Oddly, the neck differentiation in these turtles did not occur until the late Cretaceous (99–65 mya).
Reflecting their Gondwana origin, one finds only cryptodires in the Northern Hemisphere, whereas a significant number of pleurodires occur in the Southern Hemisphere. Seaturtles and soft-shelled turtles, familiar animals throughout much of the world today, appeared in the Upper Jurassic (159–144 mya); by the time the neck specializations came about in the late Cretaceous, all turtles were essentially modern.
A few very big seaturtles lived in the Mesozoic (248–65 mya) seas during the time when dinosaurs dominated the land. Archelon of the Cretaceous (144–65 mya) had a shell length of 6.3 ft (1.9 m). But the largest known turtle is a freshwater pleurodire (side-neck group) turtle with a shell length of 7.6 ft (2.3 m). Aptly named Stupendemys, this turtle was collected on a Harvard field trip to Venezuela. The animal came from Pliocene (5.3–1.8 mya) sediments, not long ago at all in terms of geologic time.
Several odd reptile clades branched off the anapsid stem, including the elephantine, terrestrial pareiasaurs, and the slim, marine mesosaurs. The pareiasaurs (among them, the well-known genus Scutosaurus) have been found in the Middle and Upper Permian (269–248 mya) of Africa, western Europe, Russia, and China. They were up to 10 ft (3 m) long and had an upright stance (unlike that of other amniotes of the time) and stout limbs that supported the massive body.
The head of pareisaurs was short and thick, with heavily sculptured bones protecting the eyes and tiny brain. The teeth had compressed, leaf-shaped crowns like those of modern leaf-eating herbivorus lizards; thus, pareiasaurs were probably the first large land herbivores.
Mesosaurs, on the other hand, were the earliest truly aquatic reptiles. These gracile animals were close to 1 yd (0.9 m) long, about a third made up of the tail. Fossil mesosaurs are known only from the adjacent coasts of southern Africa and eastern South America, reflecting the fact that Gondwana split into the two continents.
In the mesosaurs the snout was very long, and the mouth was filled with long, needlelike teeth. These teeth apparently formed a specialized straining devise that allowed the animals to feed upon small crustaceans, possibly those found in the same fossil beds as the mesosaurs. The long, compressed tail probably was used for propulsion in swimming. The posterior tail vertebrae have fracture plains, indicating that caudal autonomy (voluntary shedding of the end of the tail during stress) could have occurred; this feature may have allowed them to escape from the grasp of predators. The skull of mesosaurs originally was thought to have a temporal opening, but this was later disproved.
Euryapsida
Turning to the euryapsid reptiles, we find four highly adapted groups of sea reptiles, some of which reached ponderous proportions. Euryapsids are believed to have evolved from diapsids, having lost the lower temporal opening in the process.
They were a very important part of the marine environment during the Mesozoic and, in a sense, were as dominant in that setting as the dinosaurs were on the land. A subject that often is neglected in the course of discussions about the reasons for the demise of the dinosaurs at the end of the Cretaceous concerns the reasons behind the extinction all of the euryapsid sea reptiles at the end of the Cretaceous.
Nothosaurs lived from early to late Triassic times but were most common in the Middle Triassic (242–227 mya). They were relatively small compared with the plesiosaurs that followed them. Nothosaurs had moderately long necks and limbs modified as flippers. They are thought to have been possible ecological equivalents of modern seals and otters. Nothosaurs had sharp, conical teeth modified for catching fish. The structure of the nothosaur shoulder girdle is unique among reptiles; it provides little space for the attachment of trunk-supporting body muscles, such as occurs in land reptiles.
Plesiosaurs are thought to have evolved from nothosaurs in the Middle Triassic. They persisted until almost the end of the Cretaceous but were most abundant in the Middle Jurassic (180–159 mya) and slowly dwindled in numbers until their extinction.
Plesiosaurs had legs modified as paddles that flapped up and down like aquatic wings. This action not only pushed them through the water but also gave them a lift that allowed them to “fly” through the sea in the same manner as seaturtles and penguins.
The bones of the shoulder and hip girdles of plesiosaurs were expanded greatly below, forming an armor on the bottom of the animal that left no “soft underbelly” for attack by such predators as sharks. Two general kinds of body types were prominent in the plesiosaurs, long-necked forms and short-necked forms.
In the first group, a small head was positioned at the end of a very long neck. The body was heavy and bulbous. The teeth were conical and sharp; for this reason it is assumed that these plesiosaurs fed mostly on fishes.
The elasmosaurid clade of long-necked plesiosaurs reached a length of more than 40 ft (12.2 m) and had enormous paddles and very small heads. Some researchers have suggested that the Loch Ness monsters (if they actually existed) were long-necked plesiosaurs. If this is true, remarkable physiological changes that allowed them to adapt to icy waters must have occurred since the Mesozoic.
Short-necked plesiosaurs have practically no neck at all and a massive head. It has been suggested that they were the ecological equivalents of the killer whales of present times, as they were consummate carnivores. Kronosaurus, the largest marine reptile that ever lived, was a massive animal that reached a length of at least 42 ft (12.8 m).
This animal was found on the property of a rancher in Australia and ended up in the Museum of Comparative Zoology at Harvard. One of the problems of collecting such a large animal is how and where to exhibit it. As frequently happens, a room at the museum had to be remodeled to put this “reptilian killer whale” on exhibit.
Placodonts were marine reptiles with short, stout bodies that lived from Middle to Upper Triassic times (242–206 mya). The limbs were only moderately paddlelike. It once was suggested that placodonts were related rather closely to nothosaurs, but there is really no good evidence to support this hypothesis.
The most distinguishing feature of placodonts is the form of their teeth, which are flattened rather than pointed. In the well-known genus Placodus, the teeth along the margin of the cheek region and on the palate are large and very flat, whereas the long, narrow front teeth protrude from the end of the somewhat narrowed snout.
It is thought that Placodus used the front teeth to grasp mollusks and the hind ones to crush them. Some placodonts, such as Henodus, were superficially like turtles, in that they had an upper shell composed of numerous small polygonal bones. A lower shell was not present.
The most highly specialized marine reptiles were the ichthyosaurs, whose body took on the appearance of modern tunas, sharks, and porpoises. Ichthyosaurs lived from early Triassic to Middle Cretaceous times (248–112 mya).
The skull of ichthyosaurs is streamlined, with a long snout; the eyes are very large. The body is spindle-shaped, and the limbs are reduced to fins. The tail fin is fishlike. The individual vertebrae in the spinal cord are in the form of very short and compact biconcave discs, very similar in appearance to those of modern sharks. It is estimated that some ichthyosaurs were very active and could swim 30–40 mph (48.3–64.4 km/h). Why the ichthyosaurs became extinct in the Middle Cretaceous (ca. 121–99 mya), long before the dinosaurs and other marine reptiles, remains a mystery.
Diapsida
Aside from having two openings in the temporal region of the skull, diapsids typically have hind limbs that are longer than the forelimbs. The oldest known diapsid was a small, lizardlike animal with a body length (minus the long tail) of about 8 in (20.3 cm). This slender animal, named Petrolacosaurus, was collected from the late Pennsylvanian (ca. 303–290 mya) of Kansas.
Two distinct clades, the Lepidosauria and the Archosauria, branched off early from the diapsid trunk. These two groups are characterized by their contrasting patterns in locomotion and posture.
The lepidosaurs retained the sprawling posture and laterally directed movement of the limbs found in primitive tetrapods. Lateral undulation of the vertebral column was also an important method of locomotion for most lepidosaurs, reaching its highest degree of development in snakes.
Loosely separated skull bones, which allowed prey to be swallowed whole, was another important lepidosaur feature. On the other hand, archosaurs had limited or absent lateral undulation in the vertebral column, and the limbs were brought into a position more directly under the trunk. These modifications reached their highest degree of development in the dinosaurs and pterosaurs.
Representative species of the three groups of the Lepidosauria addressed here, the sphenodontids, lizards, and snakes, are presently alive. Turning to the sphenodontids, the tuatara (Sphenodon) of New Zealand are the only living members of this once large group. A newspaper article dating to the 1940s explained that the tuatara looked something like a lizard but really was a “living fossil.”
There are several differences between the sphenodontids and lizards, which split off from each other in the Triassic, possibly early in the epoch. In the sphenodontids, the jaw muscles are massive, which allows them to have a stronger but slower bite than lizards. Sphenodontid teeth are fused to the jaw so firmly (acrodont condition) that the jaw has a sawtooth appearance, as if the teeth are merely serrations of the bone itself.
Jaw muscles are less massive in lizards and are located farther back in the mouth, producing a weaker but faster bite. Most lizards have teeth somewhat loosely attached to the inside of the jaw (pleurodont condition), and these teeth are replaced frequently in most species.
Sphenodontids were the dominant lepidosaurs of the Jurassic (206–144 mya), but they sharply declined in the Cretaceous as the lizards began to diversify broadly. Only the two species of New Zealand tuatara have survived to the present, others having died out at the end of the Cretaceous.
The tuatara are active at much lower temperatures than most lizards, and the eggs have a gestation period of about nine months before being laid. The incubation period for the eggs is about 15 months, the longest of any known living reptile.
Growth in the young is slow, and the animals do not reach sexual maturity until they are about 20 years old. Slow growth then continues until the animal is 50 to 60 years of age. Rather than having hemipenes (double penis) like lizards and snakes, male tuatara transfer sperm to the female by an extension of the gut called the cloaca. Whereas most lizards seem to look right through a person, tuatara have a direct gaze, with big brown eyes that seem more mammalian than reptilian.
Lizards and snakes are considered to be a single clade, the Squamata (scaled reptiles). Both lizards and snakes have legless forms with a jaw structure that allows them to swallow prey whole. Snakes, however, have carried these tendencies to the extreme.
The first lizards are represented by an animal known as Paliguana, from the late Permian of South Africa. The fact that lizards had a more effective jaw structure, better hearing, and improved locomotion probably allowed them to exploit the habitats occupied by other lizardlike tetrapods, such as the sphenodontids. Most modern lizards, with the exception of the Komodo dragon (a monitor lizard that can take down deer), have not achieved large size. But in the Cretaceous, ancestral monitor lizards evolved into the ecologically important mosasaur, marine lizards that grew to 30 ft (9.1 m) in length. One giant terrestrial lizard of the past was the monitor lizard Megalania (probably 20 ft or 6.1 m long), the top predator in the Pleistocene (1.8–0.1 mya) of Australia. Fossils of Megalania at first were thought to be those of dinosaurs, but Max K. Hecht of the American Museum of Natural History proved that they were, in fact, giant lizards.
Snakes originated much later in the fossil record than lizards, at some time during the Middle to Upper Cretaceous. The four fossils that bear most closely on the ancestry of snakes are Pachyrhachis, Podophis, Lapparentophis, and Dinilysia.
The first three are from the Middle Cretaceous, but Dinilysia, the most complete and well-studied of the four, is from the late Cretaceous. The marine squamates Pachyrhachis and Podophis have been considered the most primitive snakes by some researchers, because the configuration of the skull bones resembles that of living snakes, but they have a well-developed hind-limb skeleton.
The terrestrial snake Lapparentophis, often called the “oldest snake,” is represented by vertebrae only, but they are certainly snake vertebrae, with all of the unique modifications found in generalized living snakes. Dinilysia has a skull that is a mosaic of lizard and primitive snake characters, but its vertebrae are clearly like those of a boa-like snake. Unfortunately, the whole picture of early snake evolution has been muddled by jargon-filled, convoluted arguments, the problem, as always, being the basic similarity of snakes and lizards.
Primitive snakes were dominant in the world until the Miocene, when modern snakes quickly replaced the less-advanced types. Three factors probably played a part: the return of warm and equable climates in the higher latitudes following the climatic deterioration in the Oligocene (ca. 33–23 mya), the striking spread of grassland habitats worldwide, and the evolution of many rodent groups that could be exploited by snakes as food. The largest modern snakes include the boas of the New World and the pythons of the Old World. The giant python-like snake Wonambi lived in the Pleistocene of Australia, along with the giant lizard Megalania.
Archosauria
The archosaurs, or “ruling reptiles,” branched into an impressive array of important groups, including the pseudo-suchians, crocodilians, pterosaurs, and dinosaurs. The archosaur clade may be defined by two temporal openings in the skull, one (antorbital fenestra) between the eye and the nostril and another (mandibular fenestra) in the hind part of the lower jaw.
Many of the other archosaur characters reflect skeletal changes associated with a more upright posture and front-to-back motion in the limbs. It has been pointed out by Robert Carroll that the lineages within the archosaur assemblage were all distinct from one another when they first appeared as fossils.
The living crocodilians are related to the Pseudosuchia, a diverse group of early, sometimes crocodile-like archosaurs that are linked to the true crocodiles mainly on the basis of having the so-called crocodile-normal structure of the tarsus (ankle).
The Pseudosuchia is one of the two major clades that branched from the early Archosauria. Pseudosuchians all had extensive external armor composed of bony plates. The group includes the rauisuchids, phytosaurs, and aetosaurs.
Rauisuchids are Middle and late Triassic reptiles that had a more or less upright stance and grew up to 20 ft (6.1 m) in length. Ticinosuchus, from the early part of the Middle Triassic of Switzerland, is one of the best-known forms. In the limb skeleton, the ankle and foot advanced to the level of those of modern crocodiles.
Supposedly, the upright stance developed independently of the lineage that led to the dinosaurs. Ticinosuchus had an armor of two rows of small, bony plates that extended along the trunk and a single row at the top and bottom of the tail. Sharp, piercing teeth set in sockets indicate that this animal was a carnivore.
Phytosaurs occurred abundantly in the late Triassic of North America, India, and Europe. Although they were not true crocodiles, they resembled them in body form and probably had a similar lifestyle. Phytosaurs had a very long snout, with sharp, piercing teeth in sockets along the margin of the jaws. Although the nasal openings were on top of the head, they were set far back on the snout rather than on the tip, as in true crocodiles. Both the trunk and the tail had an extensive covering of dermal armor. A variety of other contemporaneous reptiles have been found in the stomach contents of phytosaurs, thus documenting their carnivorous habits. The fossil record of phytosaurs is confined to the late Triassic.
The aetosaurs formed a distinct group also known only from the late Triassic. Rather than sharply pointed teeth, aetosaurs had small, leaf-shaped teeth that suggest a herbivorous diet. The well-known aetosaur Stagnolepus, was a bizarre
beast with short legs and an upright posture. The body was rotund and the tail massive. The body was covered with large quadrangular plates that formed extensive armor along the back, extended down the sides, covered the belly, and surrounded the tail.
The head was small in relation to the body and narrowed into a short rostrum anterior to the teeth, capped by a swollen area that indicated that the animal had a piglike nose. The relationship of aetosaurs to other archosaurian clades is poorly understood.
Crocodiles are the only surviving giant archosaurs and the top predators in many aquatic environments throughout the world today. A recent survey in Florida reported a male alligator that was 14 ft (4.3 m) long and weighed 946 lb (429 kg). Much longer crocodiles have been found in the tropics of the Southern Hemisphere.
Why crocodiles survived the rounds of extinction that occurred among the other large archosaurs is a matter of conjecture. The earliest crocodilians were terrestrial and included such forms as Gracilisuchus of the Middle Triassic of South America, which walked on two legs, and the four-legged Terrestrisuchus of the late Triassic of Europe, which had an extremely gracile body skeleton and must have been a fast runner. Terrestrisuchus was about the size of a rabbit but was a carnivore that probably scurried in and out of Triassic hiding places looking for mouse-size prey.
The mesosuchians of the Jurassic looked much more like modern crocodiles than the Triassic forms. Several of them became semiaquatic, and some invaded the marine environment.
The process of leaving land to become semiaquatic was important in terms of the body changes that took place in these transitional forms. Modern families of crocodiles, Alligatoridae (alligators and caimans), Crocodylidae (modern crocodiles), and Gavialidae (the gavial), are known to date back to the late Cretaceous. Alligatorids have a broad snout, with a relatively large number of somewhat blunt teeth.
Crocodylids have a longer, thinner snout, with a significant number of pointed teeth. Gavialids have an elongated snout and needlelike teeth, as do garfish; gavialids are consummate fish eaters. Some Cretaceous crocodiles grew to very large size and probably preyed upon dinosaurs. Sarcosuchus imperator from Africa possibly reached a length of 40 ft (12.2 m), and some from Texas were about that long as well.
Modern crocodilians were much more widespread in the world in the early Tertiary (ca. 65–34 mya) than they are today, their decline probably resulting from climatic deterioration in the Cenozoic era.
Crocodilians are the only living reptiles that give true parental care to their young, including nest guarding, helping the young exit eggs by cracking the eggs with their jaws, carrying the young from the nest to the water in their mouths, and protecting them in the water for a time. Vocal communication between the parents and the young also occurs.
The Ornithodiran evolved a neck that could be folded into an S shape and a narrow, compressed foot. One branch of the Ornithodira, the Pterosauria (flying reptiles) of the Mesozoic, had a very long, much enlarged fourth finger on the hand, which supported a wing membrane.
The other branch of the Ornithodira, which included Lagosuchus and the dinosaurs, had elongated lower legs (tibiae and metatarsals) as well as a thigh bone (femur) with the head turned inward so that it could fit into a deep socket in the hip girdle (pelvic girdle). This socket allowed the legs to support the large hip girdle, which in turn supported the body.
Pterosaurs were the first vertebrates to evolve true wing-flapping flight. They are known from the late Triassic and had become quite diverse by this time. The fact that pterosaurs had a foramen in the skull in front of the eyes (antorbital foramen), legs arranged mainly straight under the body, and a dinosaur-like ankle joint indicates their close relationship to Lagosuchus and the dinosaurs. But the fact that they had a hooked fifth metatarsal bone and an long fifth finger suggests that they diverged from the early archosaurs before the dinosaurs (which had the fifth finger reduced or lost) had split off.
Pterosaurs ranged from very small forms to by far the largest creatures that have ever flown. They are divided into two major groups, the primitive rhamphorhynchoids and the more advanced pterodactyloids. Rhamphorhyncoids are first known from the late Triassic and were dominant throughout the Jurassic.
They had a short face, a short neck, and a long tail. Some rhamphorhyncoids were as small as English sparrows. From the beginning, rhamphorhyncoids had evolved various specialized characters for flight, including a sternum (breastbone) that had a boatlike keel that supported the wing-flapping muscles, as in birds. Other birdlike bones included the scapula (shoulder blade), coracoid (upright shoulder girdle bone), and humerus (upper arm bone), all of which were specialized to contribute to active, flapping flight.
The pterodactyloids appeared in the late Jurassic and lasted until the end of the Cretaceous. These reptiles had a much longer face, longer neck, and shorter tail than the rhamphorhynchoids. The skull was highly modified in the Cretaceous genus Pteranodon, which had a long, bladelike rostrum (snout) in front of the eye and an almost equally long blade-like projection in back of the eye.
On the other hand, Pterodaustro, from the Upper Cretaceous of Argentina, had practically no skull at all behind the eye, but it did have large, elongate, upwardly curved jaws. The lower jaw had very long, closely packed teeth that are thought to have strained small invertebrate animals from the water in the same manner as baleen in whales.
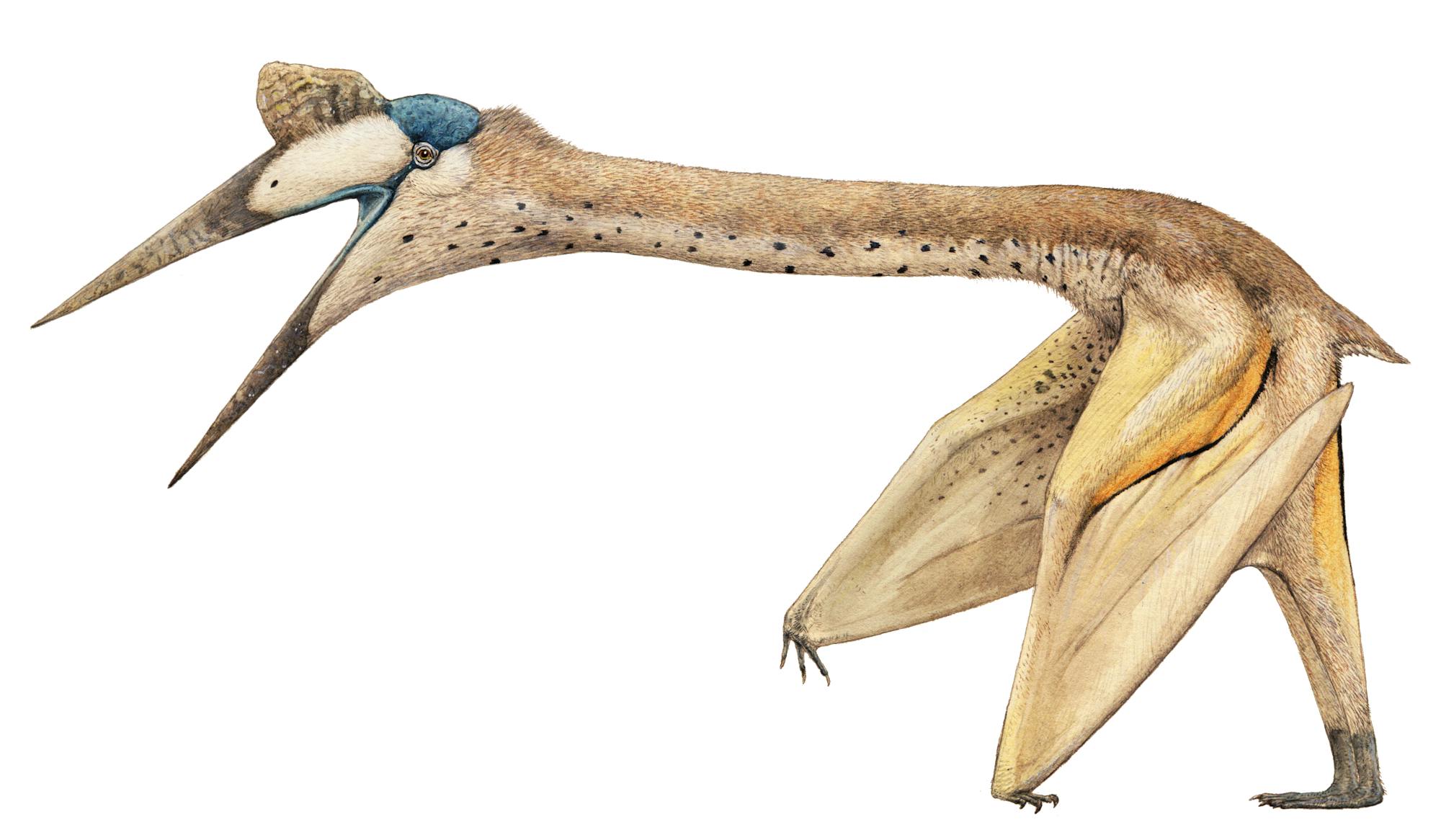
Quetzalcoatlas, from the late Cretaceous of Texas, was the largest animal ever to fly, with a wing span of more than 35 ft (10.7 m). Before the discovery of Quetzal-coatlas, British zoologist J. Z. Young suggested that Pteranodon, with its 23-ft (7-m) wing span, was probably the largest animal that could possibly fly. Robert Carroll pointed out that Quetzalcoatlas was obviously far heavier than Pteranodon.
It has been suggested that Quetzalcoatlas, might have fed upon the dead bodies of dinosaurs, like some gigantic vulture. Pterosaurs were quite different from birds, in that the wing was composed of a membrane, something like the one in bats. The difference is that in pterosaurs the membrane was supported entirely by a long, robust fourth finger. Unlike pterosaurs, bats have a wider wing membrane, and it attaches to the rear limbs.
Lagosuchus, from the Middle Triassic of South America, provides a structural link between early archosaurs and dinosaurs. Lagosuchus was only about 1 ft (0.3 m) long and had a very lightly built skeleton, with long delicate limbs; this was a humble ancestor of the gigantic animals to come.
In Lagosuchus the posterior limbs were much longer than the anterior ones. Moreover, the tibia (lower leg bone) was much longer than the femur (upper leg or thigh bone). The pelvic girdle (hip girdle) was composed of three bones forming a triradiate (three-pronged) structure. Thus, the long ilium was directed forward, the long ischium was directed backward, and the short pubis sat on top of the other two pelvic girdle bones. In the tarsus (ankle) there was a hinge between the upper and lower tarsal bones. All of these features (reflecting changes in the limbs and limb girdles, mainly the pelvic girdle and hind limbs) made Lagosuchus the most dinosaur-like of any of the primitive archosaurs.
Dinosauria
Dinosaurs fascinate more people than any other reptile group. The achievement of great size and diversity and the long domination of the earth by dinosaurs form a large part of this fascination. As is true for the great gray apparition in Mozart’s Don Giovanni or the Frankenstein monster, people love things that are terrible and wonderful at the same time. In fact, the name dinosaur comes from roots meaning “terrible lizard.”
Unique evolutionary features are evident in the hands of some dinosaurs. The joint between the thumb and the palm was structured so that when the hand was closed, the thumb bent toward the palm, indicating that the hand was used for grasping and holding objects. In effect, the bipedal gait in early archosaurs set the hands free for other functions in both pterosaurs and dinosaurs.
In the dinosaur pelvic girdle (hip girdle), the acetabulum (hip socket) either was open or was composed of cartilage in primitive forms, probably increasing the rate and range of leg motion from front to back. Also, one or more vertebrae were incorporated into the pelvic girdle, giving it more strength for vigorous locomotion.
Three main dinosaur clades are recognized by most paleontologists: the Theropoda, which were bipedal and mainly carnivorous; the Sauropodomorpha, which had long necks and were herbivorous; and the Ornithischia, which encompassed a diverse assemblage of beaked herbivores.
The earliest members of all three groups are first known at about the same time in the late Triassic. During the early part of the late Triassic, dinosaurs were an important emerging group, but they actually were dominated by pseudosuchian archosaurs and advanced synapsids (ancestral mammals with reptilian characteristics).
It was only after the great extinction that took place at the end of the Triassic that dinosaurs became the dominant group of large terrestrial vertebrates. This dominance lasted until dinosaurs became extinct at the end of the Cretaceous.
Theropods had more primitive dinosaur characters than the other two groups. For instance, theropods retained their bladelike, serrated teeth; by this feature it is known that they were carnivores. All known theropods were bipedal, and many retained grasping hands.
The most primitive known theropods were the herrerasaurids of the late Triassic of Argentina, Brazil, and North America. One specialization that these animals shared with later theropods was a joint in the lower jaw between the tooth-bearing and non-tooth-bearing portions. This innovation probably counteracted the stress associated with biting relatively large prey.
The two main clades of Theropoda are the Neotheropoda and the Coelurosauria. An early neotheropod, Coelophysis of the late Triassic, was gracile in build and had a kink in the upper jaw believed to be an adaption for holding on to small prey. Ceratosaurus of the late Jurassic was a robust form, with hornlike knobs on top of the front of its skull.
Spinosaurus of the Cretaceous had a very long snout and large, conical (rather than bladelike) teeth; the teeth at the end of the snout were larger than the rest. It has been suggested that Spinosaurus ate large fish, because some fossil localities where spinosaurs were collected contained abundant fish remains. Allosaurus of the Jurassic, a huge bipedal carnivore with a compressed head, is featured in many museums and represents a significant branch of the theropod stem called carnosaurs.

The Coelurosauria, the other main branch of the theropod stem, differed from the neotheropods in numerous ways. The brain cavity was relatively larger, and the hands were more slender. The tail tended to be very stiff.
Elements thought to be homologous to the feathers of birds, called protofeathers by some researchers, have been found in both primitive and advanced coelurosaurs, and it has been suggested that all primitive coelurosaurs may have had them. Several Jurassic and Cretaceous coelurosaurs, both small and large, were not ancestral to more advanced clades, but a clade called maniraptors became increasingly specialized.
Maniraptors are characterized by the development of a secondary palate and several changes in the structure of the brain case. They also had very slender hands and fewer tail vertebrae. Birds are a by-product of maniraptor evolution.
There are several important maniraptor groups; among them, ornithomimosaurs, such as Struthiomimus, are not bird ancestors, but they show convergence with both modern ostriches and other flightless birds. Early examples had tiny teeth, but the teeth were lost in later forms. The fingers of the hands formed a hook-and-clamp structure that may have been used to grasp branches as the animals searched for food.
Tyrannosaurids are well known in the form of Tyrannosaurus rex, popularly known as T. rex, as well as other large carnivores. It should be pointed out that tyrannosaurids evolved independently of the narrow-faced Allosaurus and its kin.
Tyarannosaurids are characterized by massive, rather than narrow heads; nipping teeth (incisors) at the front of the jaws; thickened, rather than compressed lateral teeth on the sides of the jaws; and very minimal forelimbs bearing only two claws.
These animals were fast runners for their size and by means of their jaws alone could kill their prey and render it into portions small enough to be swallowed. Early Cretaceous tyrannnosaurids were only about 10 ft (3 m) long, but later ones such as Tyrannosaurus rex were 40–50 ft (12.2–15.2 m) long with a weight of up to seven tons or more.
Other maniraptors include such groups as the oviraptorosaurs, of which Oviraptor is a well-known genus. Many species of oviraptosaurs are characterized by ornate crests and are thought to have brooded their nests in the manner of modern birds.

Dromaeosaurids include the familiar genera Deinonychus, Velociraptor, and the larger Utahraptor. These animals had long, grasping forelimbs and a large, retractable, curved claw on the second digit of the foot. Dromaeosaurids, like birds, had a backward-directed pubis and are thought by some researchers to be near the stem of bird evolution.
Found among the Sauropodomorpha are the largest land animals that ever lived; some reached a length of about 100 ft (30.5 m). All sauropods had long necks and small heads. Some of the primitive sauropodomorphs, known as prosauropods (such as Plateosaurus), spent some time on all fours, though they still could grasp objects with the hand. At the end of the Triassic, prosauropods were common large land herbivores. Long necks gave sauropodomorphs access to tree leaves, and their large body size enabled them to have a larger digestive system to process vegetation and served as a defense against small predators.
Sauropodomorphs called the Neosauropoda became a diverse group in the Jurassic and Cretaceous. Neosauropods
were characterized by columnar legs, wide hips, and specialized teeth. The two main neosauropod clades are the Diplodocoidea and the Macronaria. Diplodocoideans had thin, pencil-shaped teeth that supposedly were used for harvesting leaves and needles from the branches of trees. The skull was long and sloping, and the nostrils were positioned behind the eyes. The tails of diplodocoideans were very long and whiplike. Familiar examples are Apatosaurus (long called Brontosaurus) and Diplodocus. Shorter-necked forms include Amargasaurus of the early Cretaceous of Argentina.
In the Macronaria (“large nostrils”) clade are sauropods with enormous nostrils that are larger than the eye sockets. Quite a few members of this group had spatula-shaped teeth as well. Brachiosaurs, including the well-known genus Brachiosaurus, make up a clade that had forelimbs much longer than the hind limbs and a very long neck, a condition that brings to mind modern giraffes.
The combination of long front legs and a long neck allowed these animals to feed at the very top of trees. Sauroposeidon of the Cretaceous of Oklahoma is estimated to have been 80 ft (24.4) long, about 40 ft (12.2) of which consisted of the neck. Titanosaurs were giant macronarian sauropodomorphs that form a clade apart from the brachiosaurs. It has been discovered that advanced forms in this group developed pencil-like teeth similar to those in the diplodocids.
The Ornithischia are the third and last of the three main dinosaur clades. These animals were mainly armored, beaked, four-footed herbivores, but some of the primitive examples were at least partially bipedal. The three major clades presently recognized within the Ornithischia are the Thyreophora (armored dinosaurs), Ornithopoda (beaked, billed, and crested dinosaurs), and Marginocephalia (helmeted and horned dinosaurs).
In the diversification of the thyreophorans, one finds that the modification of armored scutes into other defensive structures is a central tendency. The relatively unarmored Scutellosaurus of the early Jurassic, thought to be a basal member of the group, had a row of bony dermal plates from the neck to the tail.
It is thought that this feature might have protected it against the small predators of the time but probably not against the large predators that appeared later in the Jurassic. Indeed, later forms modified this dermal armor into plates, spikes, and tail armor.
Stegosaurs of the late Jurassic were true armored dinosaurs, in that they had compressed plates, conical spines, or armor of intermediate shape along the middle of the back and tails that were modified as spiked clubs. Some stegosaurs had shoulder spikes, and others had masses of knobs on the throat. Ankylosaurs had an armadillo-like dermal armor fused to the head and protecting most of the body. One group of ankylosaurs had club tails.
Ornithopods may be distinguished from other Ornithischia in that the tooth row in the front portion of the upper jaw (premaxilla) is more depressed than those in the remaining upper jaw. Evolutionary tendencies in this group included an increase in size and changes in the joints of the jaw and teeth that led to a grinding mode of chewing.
Iguanodon, an early ornithopod first discovered from the early Cretaceous of England, was at one time thought to be a giant lizard, because it had leaflike serrated teeth like the modern iguana lizard. The duckbill dinosaurs of the late Cretaceous, with terminally expanded snouts, have been exceptionally well studied, and some are known literally from the cradle (nest and eggs) to the grave.
Each species of lambeosaurs had a unique crest shape and sound-producing tubes within the skull. These features provided both visual and vocal signals that indicate complex social behavior in these animals.
The last group of Ornithischia are the marginocephalians (helmeted and horned dinosaurs), represented by both bipedal and four-footed forms. The main character of this clade was a ridge or shelf of bone that overlapped the back of the skull. This group is divided into two clades, the pachycephalosaurs and the ceratopsians. Pachycephalosaurs were bipedal forms with a thick helmet of bone over the brain case; in the most derived forms, the helmet formed a thickened dome.
Many scientists believe that the dome was used for butting clashes, as occur among bighorn sheep today. Others think that because the dome was provided with a large number of blood vessels, that it might have been important in temperature regulation.
Ceratopsians had large beaks at the end of the snout. A primitive ceratopsian, Psittacosaurus, of the early Cretaceous had this deep beak but was bipedal and lacked the frill of more advanced forms. Advanced forms, called neoceratopsians, had a shelf modified as a frill at the back of the skull where the jaw muscles were attached.
Neoceratopsians had rows of teeth packed together in such a way as to provide a continuous cutting surface, but it is not known what kind of plants these animals fed on. The North American giant ceratopsians had greatly elaborated horns on the skull.
Resources – Books
Benton, Michael J. Vertebrate Paleontology. 2nd edition. Oxford: Blackwell Science, 2000.
Benton, Michael J., and D. A. T. Harper. Basic Palaeontology. London: Addison Wesley Longman, 1997.
Carroll, Robert L. Patterns and Processes in Vertebrate Evolution. Cambridge: Cambridge University Press, 1997.
Colbert, Edwin H., Michael Morales, and Eli C. Minkoff. Colbert’s Evolution of the Vertebrates. 5th edition. New York: John Wiley and Sons, Inc., 2001.
Cowan, Richard. History of Life. 3d edition. Malden, MA: Blackwell Science, 2000.
Farlow, James O., and M. K. Brett-Surman, eds. The Complete Dinosaur. Bloomington and Indianapolis: Indiana University Press, 1997.
Hallam, Arthur, and P. P. Wignall. Mass Extinctions and Their Aftermath. New York: Oxford University Press, 1997.
Holman, J. Alan. Vertebrate Life of the Past. Dubuque, IA: William C. Brown Publishers, 1994.
——. Fossil Snakes of North America: Origin, Evolution, Distribution, Paleoecology. Bloomington and Indianapolis: Indiana University Press, 2000.
Lucas, Spencer G. Dinosaurs, the Textbook. 3d edition. Dubuque, IA: William C. Brown, Publishers, 2000.
Paul, Gregory S., ed. The Scientific American Book of Dinosaurs. New York: St. Martin’s Press, 2000.
Pough, F. Harvey, Robin M. Andrews, John E. Cadle, Martha L. Crump, Alan H. Savitzky, and Kentwood D. Wells. Herpetology. 2nd edition. Upper Saddle River, NJ: Prentice Hall, 2001.
Sumida, Stuart S., and Karen L. M. Martin. Amniote Origins, Completing the Transition to Land. San Diego: Academic Press, 1997.
Weishampel, David B., Peter Dodson, and Halszka Osmólska, eds. The Dinosauria. Berkeley: University of California Press, 1990.
More information: Tiago R. Simões et al, Megaevolutionary dynamics and the timing of evolutionary innovation in reptiles, Nature Communications (2020). DOI: 10.1038/s41467-020-17190-9



{kind=link}