










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



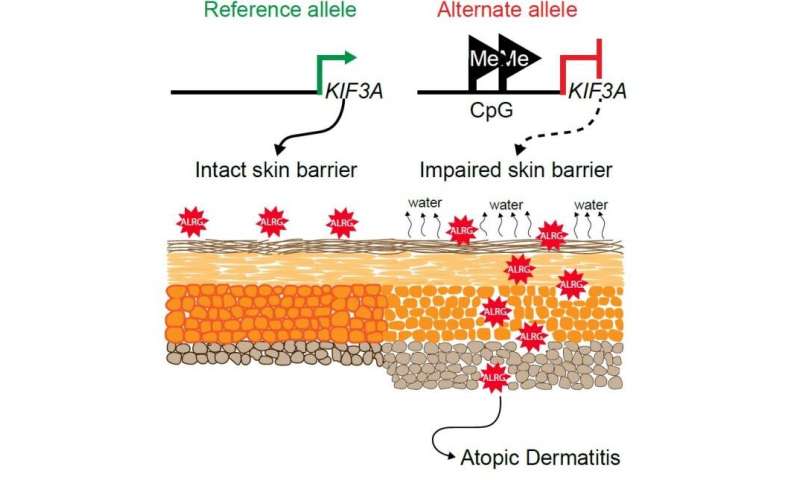
New research supported by the National Institutes of Health delineates how two relatively common variations in a gene called KIF3A are responsible for an impaired skin barrier that allows increased water loss from the skin, promoting the development of atopic dermatitis, commonly known as eczema.
This finding could lead to genetic tests that empower parents and physicians to take steps to potentially protect vulnerable infants from developing atopic dermatitis and additional allergic diseases.
Atopic dermatitis is an inflammatory skin condition that affects up to 20% of children in developed countries.
This chronic disease is characterized by dry, thickened and intensely itchy skin, particularly in skin folds.
People with eczema are more susceptible to bacterial, viral and fungal skin infections and frequently develop additional allergic diseases such as asthma.
KIF3A is a gene that codes for a protein involved in generating signals from the outside to the inside of a cell, part of a complex sensory apparatus. Previously, scientists had identified an association between two genetic variations in KIF3A and asthma in children who also had eczema.
In the new study, the researchers found that these variations, or single nucleotide polymorphisms (SNPs), changed parts of the KIF3A gene to a form that can regulate, through a process called methylation, the rate at which a gene is transcribed into the blueprint for protein production.
The investigators confirmed that skin and nasal-lining cells from people with the KIF3A SNP variants had more methylation and contained fewer blueprints for the KIF3A protein than cells in which KIF3A lacked the SNPs.
In addition, the researchers demonstrated that people with the SNP-created regulating sites had higher levels of water loss from the skin.
To determine whether lower levels of KIF3A caused atopic dermatitis, the scientists studied mice lacking the mouse version of KIF3A in skin cells.
They found that these mice also had increased water loss from the skin due to a dysfunctional skin barrier and were more likely to develop features of atopic dermatitis.
The investigators concluded that the presence of either or both of the two SNPs in human KIF3A leads to lower production of the KIF3A protein, promoting dysfunction of the barrier that normally keeps skin well hydrated, thereby increasing the likelihood that a person will develop atopic dermatitis.
Now that investigators have established that these KIF3A SNPs increase the risk for atopic dermatitis, infants could potentially be screened for them.
Therapies directed specifically at water loss from the skin, such as intensive topical moisturization regimens, could be evaluated for their ability to prevent atopic dermatitis in children with the SNPs.
Preventing atopic dermatitis in early childhood could in turn prevent a cascade of additional allergic diseases later in life, such as asthma, food allergy and allergic rhinitis – a cascade known as the atopic march.
This research was co-funded by the National Institute of Allergy and Infectious Diseases and the National Center for Advancing Translational Sciences, both part of NIH.
The study was led by Gurjit K. Khurana Hershey, M.D., Ph.D., professor of pediatrics and director of the Division of Asthma Research at Cincinnati Children’s Hospital Medical Center, which is part of the NIAID-supported Asthma and Allergic Diseases Cooperative Research Centers.
Kinesin family member 3A (KIF3A) is a susceptibility locus for both asthma1 and eczema,2, 3 suggesting that the KIF3A associations with asthma may be influenced by eczema comorbidity.
However, asthma also has a high rate of allergic rhinitis (AR) comorbidity,4 which makes it necessary to weigh the effects of both AR and eczema.
Indeed, the lack of data regarding AR was acknowledged as a limitation in a recently published meta-analysis focused on progression from eczema to asthma.5 Herein, we elucidated the associations between KIF3A single nucleotide polymorphisms (SNPs) and asthma, eczema, and AR, alone and in combination, in the Greater Cincinnati Pediatric Clinic Repository (GCPCR), a well-phenotyped biorepository of more than 7000 children, and controls from the Genomic Control Cohort (GCC), a population-based cohort of 1020 children.
Our findings were replicated in the Cincinnati Childhood Allergy and Air Pollution Study (CCAAPS), a birth cohort of 762 children with parental atopy. Details of these cohorts have been described previously.6, 7 Nonallergic controls from the GCC had no family or personal history of asthma and no personal history of any allergic disease.
The demographic and clinical characteristics for the GCPCR, GCC, and CCAAPS populations are provided in Table E1 in this article’s Online Repository at www.jacionline.org. GCPCR and CCAAPS subjects were genotyped for 19 or 7 tagging SNPs in KIF3A, respectively, as described.1
Tagging SNPs were selected from the CEU (European ancestry) and YRI (Yoruban ancestry) populations with a coefficient of determination of 0.8 and minor allele frequency of 10% or more in either population using HapMap Genome Browser release #24 (http://hapmap.ncbi.nlm.nih.gov/). Twelve of these 19 SNPs were available for the GCC control population.
All 12 SNPs were in Hardy-Weinberg equilibrium (P > .05) and had minor allele frequencies of more than 10% for GCPCR and GCC separately (see Table E2 in this article’s Online Repository at www.jacionline.org). These 12 SNPs remained in the analyses.
To assess the gene coverage of these 12 SNPs, we extracted all 42 KIF3A SNPs from Hapmap CEU (chr 5:132056221–132101163). Twenty-nine SNPs (not including the 12 tag SNPs) are in linkage disequilibrium (LD) (r2 > 0.8) with at least 1 of our 12 tag SNPs, resulting in a coverage of 96.7% using the following formula8 (G = 10 million): ((L/(R − T)) × (G − T) + T)/G.
The location and functionality of these 12 SNPs are presented in Table E2.
Most genome-wide association studies and genetic studies for asthma have not considered comorbid allergic conditions including eczema and AR. We evaluated KIF3A associations with asthma in the context of comorbid eczema and/or comorbid AR.
Comorbidity was defined as current or past disease. Of the 917 genotyped GCPCR patients for whom the genotyping passed quality control, 577 had asthma, 378 had eczema, and 622 had AR.
Among the 577 participants with asthma, 180 also had eczema and 407 had AR (see Fig E1 in this article’s Online Repository at www.jacionline.org).
The following case groups were used: all asthma, asthma + eczema, asthma without eczema, pure eczema, asthma + AR, asthma without AR, and pure AR.
Analyses were performed using GCPCR cases and GCC nonallergic control participants. We restricted the analyses to the white population because of a limited number of black participants. Assuming an additive genetic model, logistic regression was used to obtain odds ratios and P values for associations between each SNP and the respective allergic phenotypes (see Table E3 in this article’s Online Repository at www.jacionline.org).
The analyses were adjusted for age and sex. To correct for multiple testing, we determined the average pairwise LD (measured by r2 statistic) for all SNP combinations and calculated the Bonferroni correction (http://www.quantitativeskills.com/sisa/).
P values were obtained from the Wald statistic.
When all patients with asthma were compared with the nonallergic controls, 9 of the 12 KIF3A SNPs were significantly associated with asthma (Fig 1, A) as we previously published.1 When the case group was restricted to asthma + eczema, the significance of the association increased for all SNPs examined, even though the sample size decreased (Fig 1, A, and Table I). There was no association of KIF3A with the asthma without eczema group (Fig 1, A, and Table I).


We next evaluated the impact of AR. If the association of KIF3A with asthma + eczema was driven primarily by underlying AR, then the association of KIF3A with asthma + AR should be similar to asthma + eczema. However, none of the KIF3A SNPs was associated with asthma + AR (Fig 1, A). Thus, the increased association of KIF3A with asthma in the presence of comorbidity is specific for asthma + eczema.
In addition, we performed analyses using “pure” phenotypes (eg, eczema without asthma or AR and AR without asthma or eczema). Many of the GCPCR children with an eczema-only diagnosis were still of preschool age at the time of data collection, and thus unlikely to have developed asthma.
Therefore, the pure eczema group was limited to children for whom we had follow-up until at least age 7 years. Again, there was no association of KIF3A SNPs with pure eczema or pure AR, supporting that KIF3A is associated specifically with eczematous children who progress to develop asthma (Fig 1, A).
Although the SNPs genotyped herein were identified by HapMap release #24 as tagging SNPs, we found considerable LD in the region spanning KIF3A (see Fig E2 in this article’s Online Repository at www.jacionline.org). To evaluate the number of independent signals in the KIF3A region, we performed conditional analyses with rs2299007, one of the SNPs most strongly associated with asthma + eczema, as a covariate. The results gave no indication of multiple signals (Table I). Consequently, subsequent analyses were focused on rs2299007.
To further explore the specificity of eczema comorbidity among patients with asthma, we applied logistic regression model to examine the association between asthma + eczema comorbidity and KIF3A. Sex and age were included as covariates in the model.
We applied the same model to asthma + AR comorbidity. There was a significant relationship between rs2299007 and asthma + eczema (Fig 1, B), with an odds ratio of 1.6 for each additional minor allele (P = .01). The asthma + AR phenotype did not exhibit any association with KIF3A rs2299007 (P = .28).
We next replicated our findings in an independent birth cohort, CCAAPS, using the additive logistic regression model in the white population.7 Children without allergic disease or skin prick testing positivity served as controls (N = 59). P values and odds ratios were obtained after adjusting for sex. Two of the 7 genotyped KIF3A SNPs were significantly associated with the asthma + eczema phenotype, including 1 SNP, rs12186803, that was also significantly associated with asthma + eczema in GCPCR (Table I).
Using the strictest definition of genetic replication provided by Sullivan,9 we observed SNP level validation for rs12186803. In contrast, none of the SNPs was associated with asthma without eczema (P > .7). Although none of the SNPs was significant in CCAAPS after multiple testing correction, most likely because of the small sample size, all the KIF3A SNPs in the CCAAPS cohort had odds ratios of more than 2.3 for associations with asthma + eczema, which is greater than the odds ratios found in the discovery cohort.
A limitation of our study is the relatively small sample size of the subgroup with eczema only, which may in part account for the lack of association between KIF3A and pure eczema. Our results reveal that the association with asthma is evident only in the asthma + eczema group and is not seen in the asthma without eczema group. These data support a recent study that reported that the IL4/KIF3A locus is associated with asthma/eczema comorbidity.8
Although previous studies have demonstrated KIF3A genetic associations with asthma1 or eczema,2, 3 these previous studies did not characterize allergic comorbidities. Similarly, the study by Marenholz et al5 did not examine the potential role of AR. In our study, we demonstrate that the association of KIF3A is with asthma + eczema.
he presence of AR comorbidity did not increase the genetic association of KIF3A with asthma or even with asthma + eczema. Thus, our results indicate that asthma + eczema constitutes a distinct asthmatic phenotype characterized by an association with KIF3A SNPs, which is not strengthened by AR. The specific association of KIF3A with asthma + eczema may have important clinical consequences.
We acknowledge the administrative assistance of Cynthia Chappell. We thank the children and families who participated in this study.


Table E1Demographic and clinical characteristics of the 3 cohorts used in the study
| Characteristic | Discovery | Replication | |
|---|---|---|---|
| GCPCR | GCC∗ | CCAAPS† | |
| N | 917 | 412 | 392 |
| Age (y), mean ± SD | 7.95 ± 4.91 | 10.86 ± 4.29 | NA |
| Sex: male (%) | 58.2 | 47.6 | 54.8 |
| Asthma (%) | 62.9 | 0 | 13.8 |
| Eczema (%) | 41.2 | 0 | 37.0 |
| AR (%) | 67.8 | 0 | 19.6 |
| Nonallergic controls (%) | 0 | 100 | 15.1 |
SPT, Skin prick testing.∗ Nonallergic controls from the GCC did not have any family or personal history of asthma and no personal history of any allergic disease.† In the CCAAPS, children without allergic disease or SPT positivity were defined as nonallergic controls.
Table E2Annotation of KIF3A SNPs used in the discovery cohort
| SNP | Position on chromosome 5∗ | MAF (Asthma) | MAF (GCC) | HWE† (Asthma) | HWE† (GCC) | Functionality | CpG-SNP |
|---|---|---|---|---|---|---|---|
| rs9784600 | 132734155 | 0.3024 | 0.2633 | 0.8432 | 0.4457 | Intronic | |
| rs9784675 | 132734047 | 0.171 | 0.1229 | 0.7684 | 0.8181 | Intronic | |
| rs11740584 | 132711097 | 0.1398 | 0.1396 | 0.862 | 0.6811 | Intronic | X |
| rs7737031 | 132710898 | 0.136 | 0.1396 | 0.7224 | 0.6811 | Intronic | |
| rs17691077 | 132707659 | 0.1658 | 0.1165 | 0.651 | 0.4726 | Intronic | |
| rs2299007 | 132707340 | 0.1649 | 0.1165 | 0.6497 | 0.4726 | Intronic | X |
| rs3798130 | 132706454 | 0.1571 | 0.1153 | 1 | 0.4657 | Intronic, missense | |
| rs12186803 | 132704377 | 0.1049 | 0.1286 | 0.822 | 1 | Intronic | |
| rs1468216 | 132700560 | 0.1652 | 0.1165 | 0.6501 | 0.4726 | Intronic | X |
| rs2023822 | 132698654 | 0.299 | 0.2573 | 1 | 0.6079 | Intronic | |
| rs2237059 | 132697920 | 0.1675 | 0.1238 | 0.552 | 0.6553 | Intronic | |
| rs2023823 | 132697114 | 0.1649 | 0.125 | 0.6497 | 0.8208 | Intronic | X |
MAF, Minor allele frequency.∗ Genome Reference Consortium GRCh38.p2 primary assembly.† P values for Harvey-Weinberg equilibrium.
Table E3Associations between KIF3A SNPs and asthma phenotypes in GCPCR
| KIF3A SNP | All asthma (n = 577) | Asthma + eczema (n = 180) | Asthma without eczema (n = 392) | Pure eczema (n = 63) | Asthma + AR (n = 407) | Asthma without AR (n = 146) | Pure AR (n = 128) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OR | P value | OR | P value | OR | P value | OR | P value | OR | P value | OR | P value | OR | P value | |
| rs2023823 | 1.30 | .02 | 1.51 | .003 | 1.10 | .46 | 1.52 | .044 | 1.23 | .10 | 1.55 | .007 | 1.21 | .26 |
| rs2237059 | 1.53 | .003 | 1.84 | .0004 | 1.23 | .23 | 1.40 | .23 | 1.39 | .04 | 1.87 | .002 | 1.47 | .07 |
| rs2023822 | 1.08 | .61 | 1.06 | .76 | 1.06 | .73 | 1.36 | .22 | 1.04 | .82 | 1.23 | .30 | 1.00 | 1.00 |
| rs1468216 | 1.05 | .76 | 1.05 | .78 | 1.02 | .91 | 1.25 | .39 | 1.02 | .92 | 1.16 | .45 | 0.92 | .70 |
| rs12186803 | 1.58 | .002 | 1.90 | .0002 | 1.28 | .16 | 1.44 | .18 | 1.45 | .02 | 1.94 | .001 | 1.51 | .06 |
| rs3798130 | 1.59 | .001 | 1.92 | .0002 | 1.29 | .15 | 1.38 | .24 | 1.46 | .02 | 1.97 | .0007 | 1.50 | .06 |
| rs2299007 | 1.50 | .006 | 1.89 | .0002 | 1.19 | .33 | 1.48 | .15 | 1.44 | .02 | 1.72 | .008 | 1.46 | .08 |
| rs17691077 | 0.76 | .08 | 0.71 | .11 | 0.79 | .19 | 0.86 | .62 | 0.87 | .40 | 0.61 | .04 | 1.10 | .67 |
| rs7737031 | 1.59 | .001 | 1.90 | .0002 | 1.28 | .16 | 1.44 | .18 | 1.46 | .02 | 1.95 | .0008 | 1.41 | .11 |
| rs11740584 | 1.32 | .01 | 1.51 | .003 | 1.15 | .31 | 1.63 | .019 | 1.23 | .10 | 1.61 | .003 | 1.19 | .30 |
| rs9784675 | 1.48 | .006 | 1.77 | .0007 | 1.21 | .27 | 1.46 | .16 | 1.32 | .07 | 1.90 | .001 | 1.57 | .03 |
| rs9784600 | 1.44 | .01 | 1.68 | .002 | 1.20 | .30 | 1.36 | .26 | 1.30 | .09 | 1.81 | .003 | 1.49 | .06 |
Nonallergic controls from the GCC served as controls (n = 412). P value was obtained by Wald test. P < .0166 was considered statistically significant.OR, Odds ratio.
REFERENCE
- Butsch Kovacic M. – Biagini Myers J.M. – Wang N. – Martin L.J. – Lindsey M. – Ericksen M.B. – et al. -Identification of KIF3A as a novel candidate gene for childhood asthma using RNA expression and population allelic frequencies differences. PLoS One. 2011; 6: e23714
- Paternoster L. – Standl M. – Chen C.M. – Ramasamy A. – Bonnelykke K. – Duijts L. – et al. – Meta-analysis of genome-wide association studies identifies three new risk loci for atopic dermatitis. – Nat Genet. 2012; 44: 187-192
- Hirota T. – Takahashi A. – Kubo M. – Tsunoda T. – Tomita K. – Sakashita M. – et al. – Genome-wide association study identifies eight new susceptibility loci for atopic dermatitis in the Japanese population. – Nat Genet. 2012; 44: 1222-1226
- Ballardini N. – Kull I. – Lind T. – Hallner E. – Almqvist C. – Ostblom E. – et al. – Development and comorbidity of eczema, asthma and rhinitis to age 12: data from the BAMSE birth cohort. – Allergy. 2012; 67: 537-544
- Marenholz I. – Esparza-Gordillo J. – Ruschendorf F. – Bauerfeind A. – Strachan D.P. – Spycher B.D. – Meta-analysis identifies seven susceptibility loci involved in the atopic march. – Nat Commun. 2015; 6: 8804
- Butsch Kovacic M. – Biagini Myers J.M. – Lindsey M. – Patterson T. – Sauter S. – Ericksen M.B. – et al. – The Greater Cincinnati Pediatric Clinic Repository: a novel framework for childhood asthma and allergy research. – Pediatr Allergy Immunol Pulmonol. 2012; 25: 104-113
- LeMasters G.K. – Wilson K. – Levin L. – Biagini J. – Ryan P. – Lockey J.E. – et al. – High prevalence of aeroallergen sensitization among infants of atopic parents. – J Pediatr. 2006; 149: 505-511
- Barrett J.C. – Cardon L.R. – Evaluating coverage of genome-wide association studies. – Nat Genet. 2006; 38: 659-662
- Sullivan P.F. – Spurious genetic associations. – Biol Psychiatry. 2007; 61: 1121-1126
Journal information: Nature Communications



{kind=link}