










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Our mouths may be home to a newly discovered set of multi-tasking taste cells that, unlike most known taste cells, which detect individual tastes, are capable of detecting sour, sweet, bitter and umami stimuli.
A research team led by Kathryn Medler at the University at Buffalo reports this discovery in a study published 13th August in PLOS Genetics.
Taste buds in the mouth are critical to our survival and help us to decide whether a food is a good source of nutrients or a potential poison.
Taste buds employ three types of taste cells:
- Type I cells acts as support cells;
- Type II cells detect bitter, sweet and umami tastes; and
- Type III cells detect sour and salty flavors.
To better understand how taste cells detect and signal the presence of different tastes, the researchers used an engineered mouse model to investigate the signaling pathways that the animals use to relay taste information to the brain.
They discovered a previously unknown subset of Type III cells that were “broadly responsive” and could announce sour stimuli using one signaling pathway, and sweet, bitter and umami stimuli using another.
The idea that mammals might possess broadly responsive taste cells has been put forth by multiple lab groups, but previously, no one had isolated and identified these cells.
The researchers suspect that broadly responsive cells make a significant contribution to our ability to taste.
Their discovery provides new insight into how taste information is sent to the brain for processing, and suggests that taste buds are far more complex than we currently appreciate.
“Taste cells can be either selective or generally responsive to stimuli which is similar to the cells in the brain that process taste information,” commented author Kathryn Medler. “Future experiments will be focused on understanding how broadly responsive taste cells contribute to taste coding.”
Transduction
The taste cells are divisible into four types characterized by both morphological and molecular features and given the names Type I, Type II, Type III and Type IV (Figure 1).
Type I cells are similar in many ways to astrocytes and Type IV cells are immature cells, whereas Type II and Type III cells serve as the transducing elements for different taste qualities.

Four different morphological and molecularly distinct types of cells populate taste buds. Types II and III transduce different classes of tastes, whereas Type I cells are more glial-like. Type IV cells are the immature population, which develop into the other cell types over the span of a few days. Figure generated from data in 12.
Type II cells use G protein–coupled receptors for sweet (T1R2 + T1R3), umami (T1R1 + T1R3), or bitter (T2Rs) to initiate a transduction cascade, whereas Type III cells rely on ion channels for transduction of the ionic tastes of salty and sour.
The receptors and downstream signaling cascade for the Type II cells (sweet, umami, or bitter) have been well described since the early part of this century13 and involve a phospholipase C (PLC)-mediated cascade culminating in the activation of the Ca++-responsive channels TRPM5 and TRPM414 to depolarize the cell sufficiently to generate an action potential via voltage-gated Na+ channels (SCN2A, SCN3A, and SCN9A15).
Why axonless receptor cells should generate action potentials is of interest and is likely related to the release mechanism for neurotransmitter from Type II taste cells as described below.
Whereas early studies suggested that a single sweet taste receptor (T1R2 + T2R3) mediates all responses to sugars and sweeteners16, recent studies suggest that other mechanisms also play a role for glucose-containing sugars but not for artificial sweeteners.
Glucose transporters and the KATP channel, which are expressed in sweet-responsive (T1R3-expressing) taste cells17, are involved in cephalic phase insulin release independent of the neural signal for sweet transmitted to the nervous system18.
The exact mechanism by which activation of the taste cells evokes insulin release is unclear but may involve humoral rather than neural signals.
Sour
In 2006, Huang et al.19 showed that sour detection depended on cells expressing PKD2L1—cells subsequently identified as a subset of Type III cells20. In 2011, Horio et al. showed that PKD2L1 itself was not necessary for transduction of protons21.
Rather, transduction of sour involves permeation of H+ through an apical ion channel22 subsequently identified as OTOP16. Using PKD2L1 as a molecular identifier for sour-responsive taste cells, Liman and co-workers7 and Zhang et al.8 went on to confirm OTOP1 as the necessary transduction channel underlying sour taste.
The entering H+ ions not only directly depolarize the taste cells but also block Kir2.1 K+ channels23, thereby amplifying the depolarization of the entering H+ ions.
The resulting depolarization triggers voltage-gated Na+ channels (SCN2A15) to generate action potentials that activate voltage-gated Ca++ channels triggering the release of synaptic vesicles24.
In keeping with the PKD2L1 cells being the sour-transducing cells, optogenetic driving of these cells evokes an aversive response25. Curiously, another study26 reported that optogenetic driving of the PKD2L1 population drives drinking behavior in thirsty mice.
Why the mice should respond with drinking to a sensation of sour is still unresolved, although Zocchi et al.26 suggest that a subset of PKD2L1-expressing cells may convey a specific “water taste” as is known for insects27–29.
Salt
Historically, responsiveness to salt has been separated into amiloride-sensitive (AS) and amiloride-insensitive (AI) modalities30. Confounding our understanding of salt taste is that low concentrations of salt are appetitive whereas high concentrations are aversive.
Furthermore, while Na+ is important perceptually for salt, other substances, not containing Na+, also are salty-tasting. The multiple perceptual and chemical properties suggest that more than one transduction mechanism may be involved.
Supporting this concept are the results from31,32. These studies argue that the molecular correlate of AS salt, the epithelial sodium channel (ENaC), underlies the appetitive qualities of Na+ but that AI salt detection of high concentrations of Na+ relies on a subpopulation of the bitter-responsive Type II cells and a subset of the sour-responsive Type III cells5,32,33.
Furthermore, a recent study34 suggests that the AI salt transduction mechanism may directly involve Cl−, but the actual mechanism remains elusive since known Cl− channel and transporter blockers have no effect on salt taste.
Two recent works confirm a previous study35 suggesting that AS-responsive taste cells fall into a unique class of taste cells not identified by the canonical reporters (TRPM5 for Type II and CAR4 for Type III32 or PIRT for Types II and III34).
Further confounding the interpretation of these studies on Na+ transduction is the finding that, compared with rodents, humans—who enjoy low salt and avoid high salt—do not have a large AS component of salt taste36, although chimpanzees do have a minor AS component37.
Peripheral neurotransmission
Whatever means are used for transduction, the Type II and Type III cells ultimately must release one or more neurotransmitters to activate the afferent nerves.
Activity-dependent release of ATP from Type II cells38,39 and 5-HT from Type III cells40 was clearly demonstrated by several investigators, but whether other transmitters (for example, glutamate or acetylcholine) also may be involved remains unclear.
Although ATP acting on neural P2X receptors was identified as a crucial transmitter for all tastes over a decade ago41, only recently has the contribution of 5-HT acting on neural 5-HT3 receptors been elucidated in terms of transmission of sour taste42. In addition, taste cells may directly release peptides such as glucagon-like peptide 1 (GLP-1)43,44.
These additional transmitters may act to modulate adjacent taste cells45 and activate afferent nerve fibers.
Although both Type II and Type III cells require action potentials for transmitter release, the mechanisms of release for these two cell types are quite different (Figure 2).
Type III cells use a conventional synapse involving voltage-gated Ca++ channels and SNARE mechanisms to effect release of synaptic vesicles24,46,47. In contrast, Type II taste cells (transducing bitter, sweet, or umami) rely on action potentials to trigger the voltage-gated large-pore channel CALHM1 to release ATP48–50.
The pore size of this channel is sufficient to permit passage of ATP which serves as an obligatory transmitter in this system41,51,52.
The biophysical properties of CALHM1 as described in the seminal article on this channel48 did not exactly match the properties of the release channel in taste buds.
A more recent report49 showed that the channel in taste buds consists of two subunits, CALHM1 and CALHM3, which together form a channel matching the properties of the Type II cell release channel.
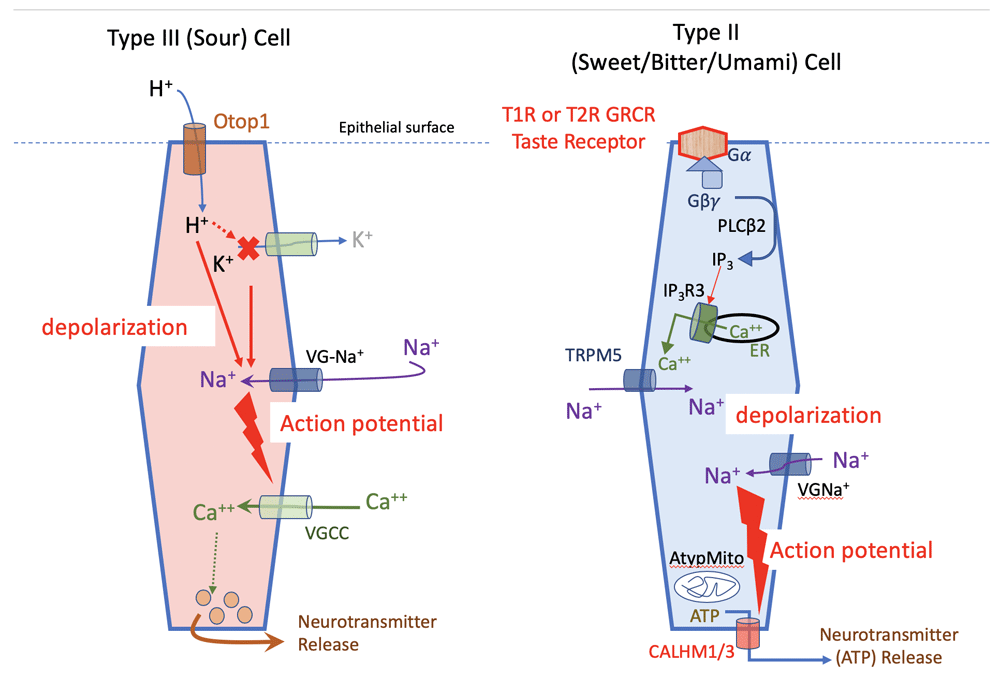
Transduction pathways for the two different types of taste-transducing cells: Type III for sour and Type II for sweet, bitter, or umami. The different responsiveness of Type II cells is dictated by the type of receptor each cell expresses, not by downstream members of the transduction cascade. AtypMito, atypical mitochondria; ER, endoplasmic reticulum; Gα, alpha subunit of G protein; Gβγ, beta-gamma subunits of G protein; IP3, inositol trisphosphate; IP3R3, inositol trisphosphate receptor isoform 3; PLCβ2, phospholipase C isoform β2; TRPM5, transient receptor potential cation channel subfamily M member 5; VGCC, voltage-gated calcium channel; VG-Na+, voltage-gated sodium channel.
Whereas the mechanism of release for ATP is well established for Type II cells, the source of ATP for transmission of sour information from Type III cells remains enigmatic.
No one has yet demonstrated direct release of ATP from Type III cells, yet transmission of Type III cell taste qualities (for example, sour) is dependent on intact purinergic signaling to P2X receptors41.
Type III cells do not express CALHM148 and so do not use CALHM1/3-mediated release. Another possible means of release of ATP is via synaptic vesicles, but Type III cells are reported to lack the vesicular ATP-transporter, VNUT53, presumed to be necessary for loading of synaptic vesicles with ATP. So, what other possible sources exist for release of ATP?
One suggestion is that Type III cells may trigger ATP release via interaction with other taste cell types. If so, this interaction does not require participation of Type II cells since mice lacking Type II cells (Skn1A-KO)54 and CALHM1 KO48 mice show nearly normal responses to sour.
Hence, the transmission of sour information to taste nerves does not require the presence of Type II taste cells nor the function of CALHM1 to release ATP.
Furthermore, a recent study showing high-resolution reconstructions of taste buds shows that Type III cells seldom directly contact Type II cells since processes from Type I cells intervene12, suggesting that any interactions between Type II and Type III cells may be indirect.
REFERENCE LINK : https://f1000research.com/articles/8-2117/v1
Source: PLOS
Original Research: Open access
“A subset of broadly responsive Type III taste cells contribute to the detection of bitter, sweet and umami stimuli” by Debarghya Dutta Banik, Eric D. Benfey, Laura E. Martin, Kristen E. Kay, Gregory C. Loney, Amy R. Nelson, Zachary C. Ahart, Barrett T. Kemp, Bailey R. Kemp, Ann-Marie Torregrossa, Kathryn F. Medler. PLOS Genetics.



{kind=link}