











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new tissue-transport device which was inspired by the egg-laying organs of parasitic wasps could greatly advance the field of minimally invasive surgery (MIS), according to a new study in Frontiers in Bioengineering and Biotechnology.
The prototype, developed by researchers at Delft University of Technology in the Netherlands, could allow MIS techniques to be used in previously hard-to-reach areas of the body and vastly reduce the trauma from surgery, and therefore recovery time, of patients.
The ovipositor is an ultra-thin organ, shaped like a flexible hollow needle, used by parasitic wasps to lay their eggs in trees or live hosts.
Inside this delicate organ is a series of tiny blades that join together with a tongue-and-groove mechanism.
The blades can slide independently of each other to create friction forces by using their mechanical movement, and it is these friction forces that the researchers used in the implementation of their design.
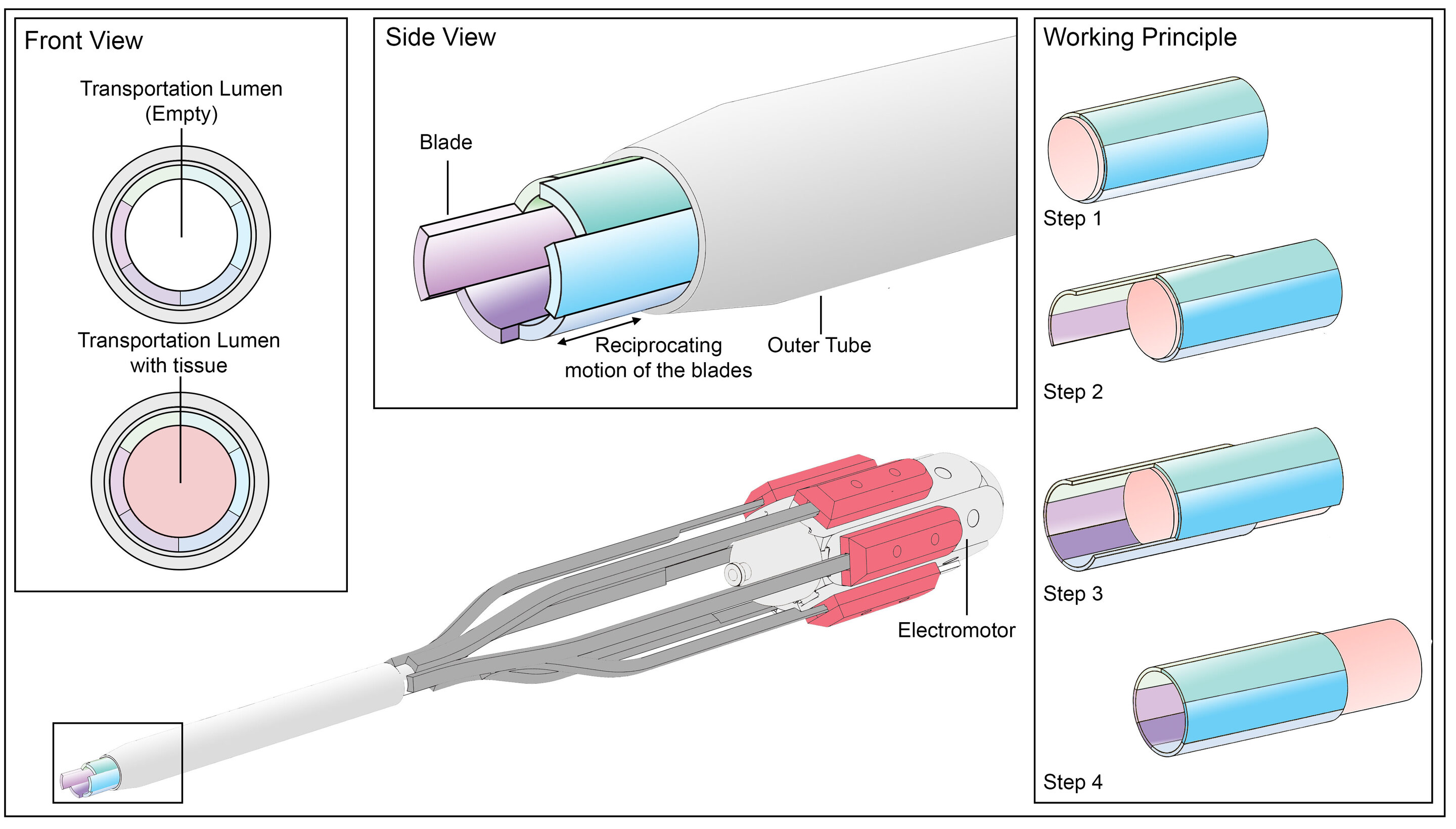
“The wasp ovipositor is so thin that it can’t actually fit any muscles within it, so we knew it was a clever mechanical solution worth studying to see if we could recreate it,” explains lead author Dr. Aimée Sakes of the Bio-Inspired Technology Group at Delft University of Technology, led by Prof Dr. Paul Breedveld.
“The ovipositor-inspired transport system uses friction generated between the blades and the tissues to transport those tissues, in the same way the wasp ovipositor works to transport wasp eggs.”
Applying these friction based techniques in the design of new surgery devices may allow researchers to surpass the boundaries faced by current tools, which work by suction.
This poses challenges, as suction-based devices lose their efficacy once they are decreased in size past a certain point, making it difficult to limit the tissue damage done in surgery. Dr. Sakes highlights: “We already see challenges arise with current devices, as they often get clogged when removing things like blood clots.”
The bio-inspired tissue-transport tool in action. Credit: A. Sakes and coauthors
“We also currently can’t reach remote locations in the human body or perform surgery in miniature structures, such as parts of the brain, due to the relatively large size of MIS instruments. However, our system could potentially enable the removal of tumorous tissues deep inside the human body through miniature incisions in future,” says Dr. Sakes.
The main drawback of the newly developed tool is that the speed at which it transports tissues is still substantially slower than current suction-based devices, so further research is still needed before the system is ready to be used in surgery.
However, the research offers very promising potential for treating diseases currently not treatable and reachable with existing equipment, especially with a view to further reduction of patient trauma in MIS.
From a mechanical perspective, it is very difficult to drill into a solid substrate with a very thin probe, because it can easily bend and break. Parasitic wasps, however, do this regularly when they use their slender ovipositors to search for hosts in solid substrates, such as fruits or even wood (1⇓–3).
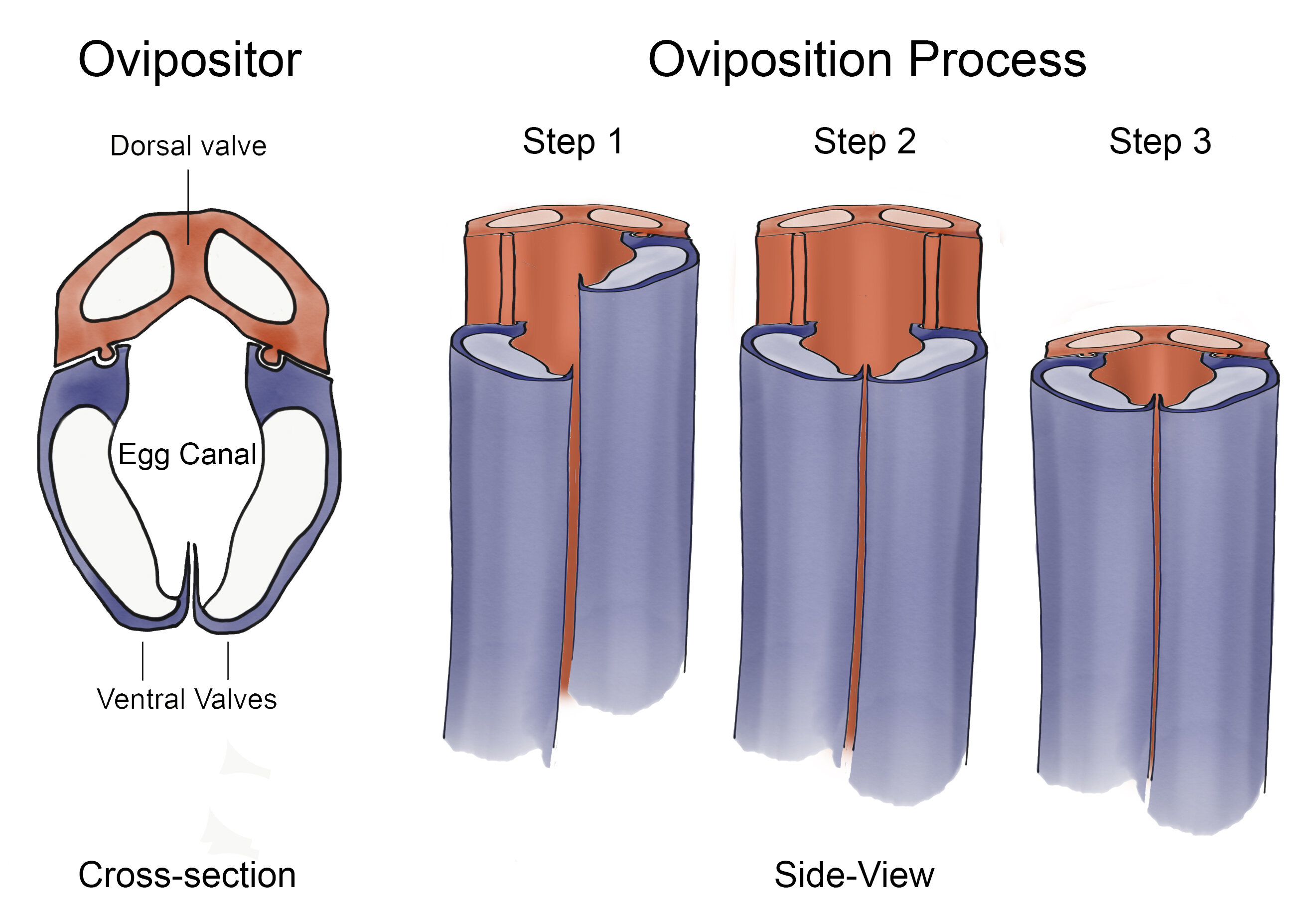
The general morphology of the ovipositor is similar across all wasp species (4, 5); it consists of four elements, called valves, of which two are often merged such that three functional valves remain (Fig. 1).
In most species, the distal part of the ovipositor is morphologically distinct (3, 6), which we will refer to as the tip. The valves can slide along each other (5, 7) and do not get dislocated under natural conditions, because they are longitudinally connected via a tongue-and-groove mechanism (5, 8⇓–10).
The ovipositor and the “wasp waist,” a constriction of the body between the first and second abdominal segment (11), are essential in probing behavior and are therefore considered to be instrumental in the evolution of the order (11–15).
The shape, structure, and mechanical properties of the ovipositors are putatively adapted to the substrates into which the animals need to probe (6, 16–18), and because both substrates and hosts are so diverse, this might have resulted in high species diversification of the hymenopterans (13, 14).
However, to understand the observed diversity in the ovipositor shapes, understanding of the probing mechanics is essential.

μm (A) and 10 μ
μm (B, Inset)].
Wasps are faced with two problems when searching for hosts in (solid) substrates: (i) how to insert the ovipositor without buckling/breaking it and (ii) how to maneuver with the ovipositor to reach the target.
Buckling is a mechanical failure of a structure which occurs, for instance, when a beam cannot withstand the applied axial load and bends, possibly beyond its breaking point. As buckling occurs more easily in slender beams, this is a real danger for parasitic wasps.
Buckling depends on four parameters: (i) the axial load applied on the beam, (ii) the second moment of area of the beam, (iii) how well is the beam fixed on both ends (i.e., “free to slide sideways,” “hinged,” or “fixed”), and (iv) the length of the beam.
During puncturing, axial loading of the ovipositor cannot be avoided, so only the other factors can be adjusted. The second moment of area is largely determined by the diameter of the ovipositor and its wall thickness.
To simplify insertion, the ovipositor must be as thin as possible, while the internal channel needs to be big enough for an egg to pass. Both of these requirements increase the chance of buckling. In all wasps, the ovipositor is fixed internally to the reproductive system and the muscles that move the ovipositor (4, 19), so very little variance can be expected related to the fixation of the ovipositor.
A parameter that can be changed is the “functional” length of the ovipositor. Some wasps protrude only a small part of the total ovipositor outside their bodies before puncturing the substrate. The part retained in the abdomen is then either strongly coiled or telescopically retracted (20, 21).
In other species, the functional length of the ovipositor is reduced by supporting it by clamping the ovipositor with parts of their hind legs (11, 18, 22) or with specialized sheaths (1, 10, 23, 24).
Little is known about the mechanisms parasitic wasps use for further insertion and buckling prevention of the ovipositor after the initial puncturing of the substrate. Vincent and King (23) hypothesized a mechanism that wasps might use based on the morphology of the ovipositor (Fig. 2A).
In the proposed mechanism, wasps apply a pulling force on two of the three valves, which are kept stationary because of the hook-like structures on their tips function as anchors. These valves serve as guides for the third valve that is pushed inward. According to the hypothesis, buckling of the protracted valve is avoided by limiting the amplitudes of forward motion.
By alternating the protraction and retraction of the valves, the ovipositor is further inserted into the substrate, while avoiding excessive net push forces and axial loads that could damage the ovipositor (23).

Hypothesised insertion and steering mechanisms in 2D. Full arrows represent push forces and empty arrows pull forces. (A) The push–pull mechanisms (23) (only two valves are shown for clarity). Inner friction (Fin) is considered negligible. Friction along the shaft (Ff) of the two pulling valves, together with the hook forces (Fh), keep the ovipositor anchored in the substrate and counteract the friction and hook (cutting) forces of the pushing valve (modified from ref. 23). (B) A bevel shape of a needle leads to bending due to asymmetrical tip forces (Ftip). (C) Restriction in inter-element displacements (32) causes bending due to tensile (gray arrows) and compression (small black arrows) forces (modified from ref. 32). (D) Arched ovipositors bend due to differential sclerotization of valve segments (34); see text (modified from ref. 34). (E) Pretension of individual elements (35) leads to in-curving upon their protraction as observed in hemipteran mouthparts (modified from ref. 35).
The second challenge during oviposition is that the wasps need to steer the ovipositor tip in the direction of the desired target (25). To do so, the ovipositor needs flexibility and a steering/bending mechanism that adjusts the tip direction during probing. Proposed bending mechanisms can be divided into passive and active ones (Fig. 2 B–E). Passive bending originates from mechanical interactions of the inserted ovipositor with the substrate. Active bending occurs when bending moments originate from the relative movements of the ovipositor valves.
Passive bending presumably occurs when an ovipositor has an asymmetric beveled tip (Fig. 2B). The asymmetric forces acting on such a tip push the tip away from a straight path (26, 27). Rotation of the bevel can be used to adjust the tip direction during insertion (28).
The tips of most ovipositors across species are asymmetric (6, 29, 30) and can thus potentially function as a bevel. The bevel shape can presumably be enhanced by changing the relative positions of the valves. An adjustable bevel may control the degree of bending, similar to what has been proposed for a new generation of steerable needles (31).
Three mechanisms have been proposed for active bending. In the first mechanism, special anatomical structures limit the motion range of individual valves (32, 33). Bending occurs due to tension and compression in individual valves (Fig. 2C). A second active bending mechanism relies on differences in valve sclerotization (34).
The distal part of ovipositors relying on this bending mechanism consists of heavily sclerotized, stiff arches, alternated with less sclerotized and flexible nodes. At rest, the arches and nodes of the dorsal and ventral valves are aligned and the ovipositor is approximately straight. When the ventral or dorsal valves are protracted, the arches align with nodes, which leads to bending (Fig. 2D).
The third possible mechanism of active bending has been hypothesized for the control of hemipteran mouthparts. Similar to the ovipositors, the hemipteran mouthparts consist of multiple slender elements that are interconnected longitudinally and are able to slide along each other. It is assumed that bending moments in hemipteran mouthparts originate from (pre)tension of the elements (35, 36).
The elements possess a certain level of inner tension and tend to curve to one side when not opposed. At rest, the elements are aligned with their tips so they counteract each other, resulting in a straight structure. When an individual element protracts, its tip curves inward toward the other elements (Fig. 2E).
In all three mechanisms, the amplitude of the protraction and retraction of the valves probably correlates with the amount of bending and offers a way to control the curvature of the ovipositor during insertion.
Despite the proposed importance of ovipositors for the evolutionary success of hymenopterans, there is only a small number of empirical studies quantifying mechanical properties of the ovipositors (6, 16⇓–18).
The proposed theories of probing are based on morphological data, with only a few studies focusing on the ovipositor inside the substrate (25, 37), but no one has ever analyzed the dynamics of probing inside the substrate. In this work, we aim to quantify the ovipositor use (range, speed, and curvature of probing) in relation to substrate density and to determine which of the proposed methods of insertion and steering are used by parasitic wasps.
We do this using the species Diachasmimorpha longicaudata, which provides an excellent example because of its long and slender ovipositor. Extrapolation of our results will also provide insight into probing and steering possibilities of other groups of parasitic hymenopterans and possibly of hemipterans and mosquitoes, as they use similar structures to probe for food.
In addition, our study will add to the understanding of the functional demands acting on the ovipositor and the mechanism for drilling with slender probes. This, in turn, can be applied in the development of man-made instruments for tunneling, insertion, or probing. Such knowledge will presumably also help in the development of novel steerable surgical tools (38–43).
reference link: https://www.pnas.org/content/early/2017/08/23/1706162114
More information: Aimée Sakes et al, Development of a Novel Wasp-Inspired Friction-Based Tissue Transportation Device, Frontiers in Bioengineering and Biotechnology (2020). DOI: 10.3389/fbioe.2020.575007



{kind=link}