











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A newly identified genetic factor allows adult skin to repair itself like the skin of a newborn babe.
The discovery by Washington State University researchers has implications for better skin wound treatment as well as preventing some of the aging process in skin.
In a study, published in the journal eLife on Sept. 29, the researchers identified a factor that acts like a molecular switch in the skin of baby mice that controls the formation of hair follicles as they develop during the first week of life.
The switch is mostly turned off after skin forms and remains off in adult tissue.
When it was activated in specialized cells in adult mice, their skin was able to heal wounds without scarring. The reformed skin even included fur and could make goose bumps, an ability that is lost in adult human scars.
“We were able to take the innate ability of young, neonatal skin to regenerate and transfer that ability to old skin,” said Driskell, an assistant professor in WSU’s School of Molecular Biosciences.
“We have shown in principle that this kind of regeneration is possible.”
Mammals are not known for their regenerative abilities compared to other organisms, such as salamanders that can regrow entire limbs and regenerate their skin.
The WSU study suggests that the secret to human regeneration might be found by studying our own early development.
“We can still look to other organisms for inspiration, but we can also learn about regeneration by looking at ourselves,” said Driskell. “We do generate new tissue, once in our life, as we are growing.”
Driskell’s team used a new technique called single cell RNA sequencing to compare genes and cells in developing and adult skin. In developing skin, they found a transcription factor – proteins that bind to DNA and can influence whether genes are turned on or off.
The factor the researchers identified, called Lef1, was associated with papillary fibroblasts which are developing cells in the papillary dermis, a layer of skin just below the surface that gives skin its tension and youthful appearance.
When the WSU researchers activated the Lef1 factor in specialized compartments of adult mouse skin, it enhanced the skins’ ability to regenerate wounds with reduced scarring, even growing new hair follicles that could make goose bumps.
Driskell first got the idea to look at early stages of mammalian life for the capacity to repair skin after learning of the work of Dr. Michael Longaker of Stanford University. When performing emergency life-saving surgery in utero, Longaker and his colleagues observed that when those babies were born they did not have any scars from the surgery.
A lot of work still needs to be done before this latest discovery in mice can be applied to human skin, Driskell said, but this is a foundational advance.
With the support from a new grant from the National Institutes of Health, the WSU research team will continue working to understand how Lef1 and other factors work to repair skin.
Also to help further this research, the Driskell lab has created an open, searchable web resource for the RNA sequence data for other scientists to access at skinregeneration.org.
Understanding how to induce skin regeneration instead of scarring will have broad implications clinically and cosmetically (Walmsley et al., 2015b). One of the main characteristics of scars is the absence of hair follicles, indicating that their regeneration in a wound may be a critical step in achieving scar-less skin repair (Yang and Cotsarelis, 2010).
Interestingly, human embryonic skin has the capacity to regenerate without scars (Lo et al., 2012). Similarly, neonatal and adult mouse skin has the capacity to regenerate small non-functional hair follicles under specific conditions (Figure 1c-d)(Ito et al., 2007; Rognoni et al., 2016; Telerman et al., 2017).
These insights have prompted efforts in the field to define the molecular triggers that promote hair development in skin, with the ultimate goal of devising a way to regenerate fully functional hairs in adult skin wounds as a therapeutic modality (Yang and Cotsarelis, 2010).

Human and mouse skin are similar in their overall structural complexity, indicating that mouse skin can be a useful model to study skin development and wound repair (Chen et al., 2013). Murine hair follicle and skin development primarily occurs between embryonic day 12.5 (E12.5) to post-natal day 21 (P21) (Muller-Rover et al., 2001).
During this time different fibroblast lineages are established that respond to the changes in the environment to support hair follicle and skin development (Driskell et al., 2013; Jiang et al., 2018; Rinkevich et al., 2015; Rognoni et al., 2016).
Fibroblasts that support hair follicle development differentiate from the papillary fibroblast lineage, into dermal papilla (DP), dermal sheath (DS), and arrector pili (AP) cells (Driskell et al., 2013; Plikus et al., 2017). Reticular fibroblasts, which cannot differentiate into papillary fibroblast lineages, secrete Extra-Cellular-Matrix and form into adipocytes (Driskell et al., 2013; Schmidt and Horsley, 2013).
By post-natal day 2 (P2) fibroblast heterogeneity is fully established with the presence of the defined layers of the dermis (Figure 1a)(Driskell et al., 2013).
Skin maturation occurs after P2 with the formation of the AP and the completion of the first hair follicle cycle, which results in the loss of a defined papillary fibroblast layer (Figure 1b) (Driskell et al., 2013; Rognoni et al., 2016; Salzer et al., 2018).
We have previously shown that papillary fibroblasts are the primary source of de-novo dermal papilla during skin development, which are required for hair formation (Driskell et al., 2013).
Furthermore, it has been suggested that adult murine skin form scars due to the lack of a defined papillary layer (Driskell et al., 2013; Driskell and Watt, 2015). Consequently, expanding this layer in adult skin might support skin regeneration in adult mice.
The use of scRNA-seq in the murine skin has established useful cell atlases of the skin during development and homeostasis (Guerrero-Juarez et al., 2019; Haensel et al., 2020; Joost et al., 2020; Joost et al., 2018; Joost et al., 2016; Mok et al., 2019). In addition, scRNA-seq studies investigating wound healing have so far focused on comparing scarring, non-scarring, or regenerating conditions (Guerrero-Juarez et al., 2019; Haensel et al., 2020; Joost et al., 2018).
These studies have helped to identified key markers of the newly discovered skin fascia, such as Gpx3, which recently has been shown to contribute to scar formation (Correa-Gallegos et al., 2019; Grachtchouk et al., 2000; Joost et al., 2020). These, scRNA-seq studies have revealed that transgenic activation of the Shh pathway in the alpha-smooth-actin cells in scarring wounds, which includes pericytes, blood vessels, and myofibroblasts, can support small non-functional hair regeneration (Lim et al., 2018).
However, activation of Shh pathway in dermal fibroblasts is associated with malignant phenotypes and will perturb development and homeostasis such that it may not be a safe target to support human skin regeneration clinically (Fan et al., 1997; Grachtchouk et al., 2000; Oro et al., 1997; Sun et al., 2020).
Altogether, these findings suggest that an overall comparison of developing, homeostatic, scarring, and regenerating skin conditions will yield important discoveries for molecular factors that can safely support skin regeneration without harmful side effects.
The Wnt signaling pathway is involved in regulating development, wound healing, disease and cancer (Nusse and Clevers, 2017). Studies that activated Wnt and β-catenin in skin have led to important discoveries for wound healing but have produced contrasting results in the context of fibroblast biology and hair follicle formation (Chen et al., 2012; Enshell-Seijffers et al., 2010; Hamburg-Shields et al., 2015; Lim et al., 2018; Rognoni et al., 2016).
Wnt is a secreted protein that activates a cascade of events that stabilizes nuclear β-catenin, which operates as a powerful co-transcription factor of gene expression. Importantly, β-catenin along cannot activate the expression of Wnt target genes without co-transcription factors.
There are four Wnt co-transcription factors Tcf1 (Tcf7), Lef1, Tcf3 (Tcf7l2), and Tcf4 (Tcf7l2). These co-transcription factors modulate the functional outcome of Wnt signaling by binding to different target genes (Adam et al., 2018; Nguyen et al., 2009; Yu et al., 2012).
In the context of wound healing and regeneration, it is not known how differential expression of Tcf/Lef can modulate fibroblast activity via Wnt signaling.
Since it has been shown that embryonic and neonatal skin have the potential to regenerate hair follicles upon wounding (Hu et al., 2018; Rognoni et al., 2016), we set out to identify the cell types and molecular factors that define this ability in order to transfer this regenerative potential to adult tissue. Our work has identified Lef1, as the factor in fibroblast of developing skin, that can transform adult tissue to regenerate without harmful phenotypes.
Neonatal skin regeneration requires Lef1 expression in fibroblasts
Based on our scRNA-seq findings, we hypothesized that Lef1 defines a neonatal papillary fibroblast that supports skin regeneration during neonatal wound healing. T
o test if Lef1 in fibroblasts is required to support neonatal regeneration, we produced a tissue specific knockout model. We utilized the fibroblast specific promoter Dermo1/Twist2 to drive Cre expression, bred with a mouse line with flox sites flanking Exon 7 and 8 of the Lef1 locus (Yu et al., 2012).
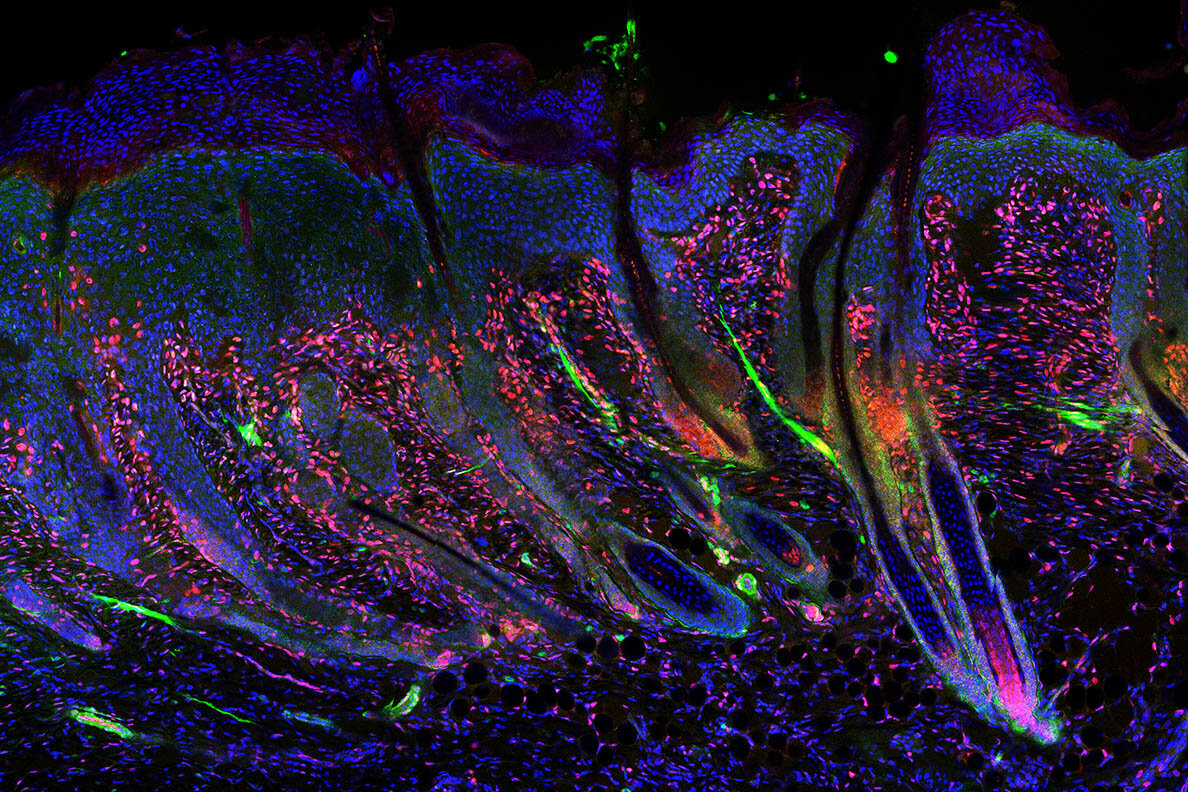
We called this mouse line DermLef1KO (Figure 3a). DermLef1KO mice were viable and fertile with small shifts in hair follicle phenotypes that resulted in less dense fur (Manuscript in Prep). We confirmed dermal Lef1 ablation from the papillary fibroblast at P2 and from adult DP by immunostaining (Figure 3b-e).
We also performed 2mm wounds on P2 DermLef1KO and wild type littermates harvesting at 7dpw (Figure 3f). Our analyses revealed that Lef1 was expressed in the de-novo DP and regenerating hair follicle buds of wild type mice (Figure 3g,i), but that wounded DermLef1KO mice lacked regeneration (Figure 3h,j,k). We conclude that Lef1 expression in neonatal fibroblasts is required for hair follicle regeneration in wounds.

reference link : https://www.biorxiv.org/content/10.1101/2020.06.11.147066v3.full
More information: Quan M Phan et al, Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds, eLife (2020). DOI: 10.7554/eLife.60066



{kind=link}