










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


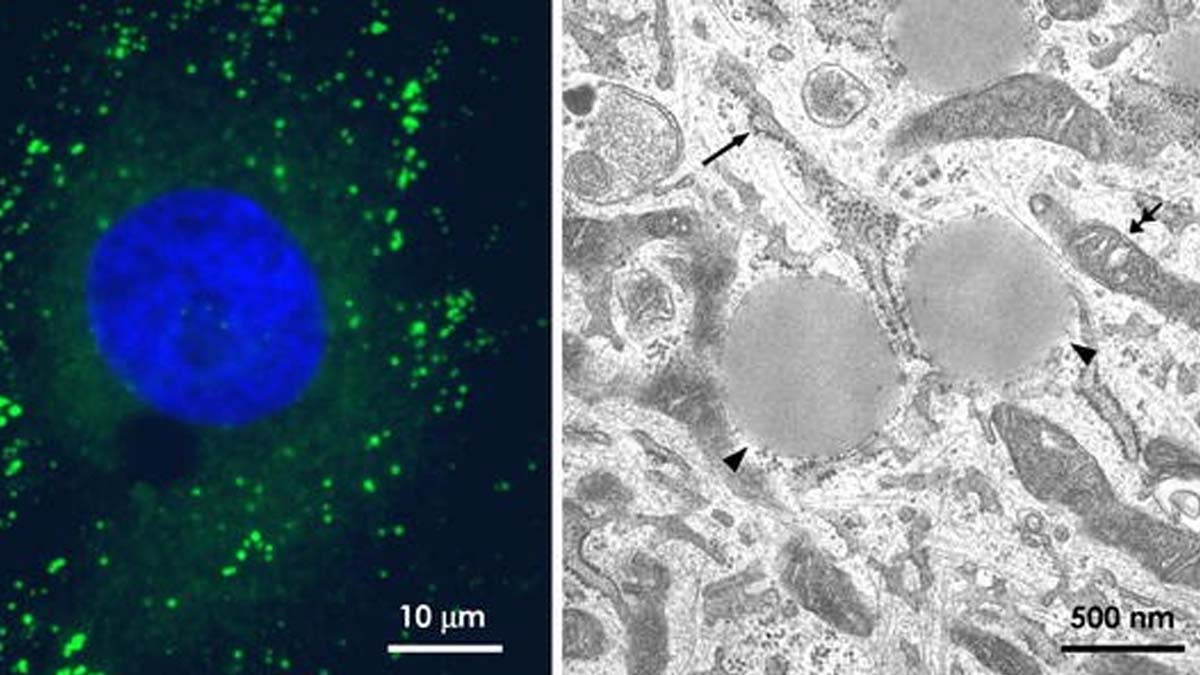
A study published in the journal Science and coordinated by researchers of IDIBAPS, the UB and teh Spanish National Center of Cardiovascular Research (CNIC) describes a new immune defense mechanism unknown until now.
It is a mechanism orchestrated by lipid droplets (LDs), the cellular organelles capable of attracting and eliminating invading pathogens.
Researchers from Spain, the United States, Australia and Brazil have participated in the international project, financed by the Human Frontier Science Program, with the collaboration of scientists from the CNIC and IIBB-CSIC. Albert Pol, ICREA professor at IDIBAPS, where he leads the Lipid Trafficking and Diseases team and associate professor at the UB School of Medicine and Health Sciences, and Robert G. Parton, from the Institute of Molecular Biosciences at the University of Queensland, are the study coordinators.
The first authors of the work are Marta Bosch, a researcher from the IDIBAPS group, and Miguel Sánchez-Álvarez, from the CNIC Mechanoadaptation and Caveolas Biology group.
LDs are the organelles where our cells accumulate nutrients that, in the form of fat, provide the necessary energy for them to develop their function.
For example, LDs provide the energy for the heart to beat, the liver to do its metabolic function, or the muscle to move.
“The lipid droplet is like the pantry of our cells, where we accumulate the food that we will use later. This happens in all eukaryotic cells, from yeasts or insects to plants or mammals,” says Albert Pol.
When viruses or bacteria infect the host cell, they need great amounts of nutrients to multiply and to get them to reach the LD. In the study published in Science the researchers have shown that, in response to infection, DLs organize complexes of antibiotic and antiviral proteins that act cooperatively to fight the pathogen and eliminate it.
It is a mechanism that would work in all cells of the body, not just professional cells of the immune system such as macrophages. This defense strategy has also been observed in insects, suggesting its importance during the evolution of our innate immunity.
The key to the innate immunity of cells
Researchers have shown that to protect themselves from infection, cells place large amounts of antibiotics and antiviral proteins on LDs.
In total, comparing the surface area of DL in normal cells and infected cells, the study has identified 400 candidates that would perform the protective function of LDs when they are in contact with the pathogen.
“In this study, we have focused on six of these proteins and we have shown that they protect the cell during the infection of different types of bacteria” explains Marta Bosch.
“The concentration of these antibiotic and antiviral proteins in a single compartment inside the cell allows creating synergies while reducing their toxicity and allowing the rest of the cellular machinery to function normally,” says Miguel Sánchez-Álvarez.
Furthermore, the study shows that this strategy allows a broad-spectrum response, that is, there are many antibiotics and antivirals with different mechanisms of action.
It also allows the generation of cooperative mechanisms to attack the infection. “There are synergies between proteins, and, for example, one breaks the membrane of the pathogen and the other destroys its genomic material,” authors explain.
“This study represents a paradigm shift since until now it was thought that LDs were at the service of viruses or bacteria during infection,” says Albert Pol. “Given the widespread resistance to current antibiotics, this study has deciphered an important defense mechanism that could be used for the development of new therapeutic strategies to stop infections,” he concludes.

Lipid droplets (LDs) are storage organelles that can modulate lipid and energy homeostasis, and historically, this was considered their defining role. More recently, LDs have emerged as a dynamic organelle that frequently interact with other organelles and are involved in protein sequestration and transfer between organelles.
LDs have also been demonstrated to act as a scaffolding platform to regulate signalling cascades, highlighting their diverse functions1–4.
The role of LDs in an infection setting has not been well studied, however, it has been demonstrated that LDs accumulate in leukocytes during inflammatory processes, and they are also induced in human macrophages during bacterial infections2.
Multiple bacterial strains, including Mycobacterium spp., Chlamydia spp., Klebsiella spp. and Staphylococcus spp. are known to upregulate LDs very early following bacterial infection in both primary and cell culture macrophage models, and this has also been seen for a number of bacterial species in rodent macrophage cell lines5–7.
Interestingly, Trypanosoma cruzi infection of macrophages also induces LDs, however, this response takes 6-12 days to occur following infection8.
Bacterial-induced LD induction in immune cells has been shown to depend on toll-like receptor engagement, mainly via TLR2 and TLR4, however, the role of LDs in the outcome of bacterial infection remains largely unknown, and the exact mechanisms for controlling LD induction remain elusive9,10. It has been suggested in recent work in the zebrafish model that embryos with higher levels of LDs are more protected from bacterial infections11 and work in the Drosophila embryo has demonstrated that LDs can bind to histones which are released upon detection of intracellular bacterial LPS and act in a bactericidal manner12
Interestingly, LD induction has been demonstrated to be a direct result of immune activation of macrophages by IFN-y in a HIF-1 dependent signalling pathway13. M. tuberculosis acquires host lipids in the absence of LDs under normal conditions, however, IFN-y stimulation of macrophages results in redistribution of host lipids into LDs where M. tuberculosis is unable to acquire them13.
IFN-y induced LDs have also been shown to enhance expression of genes involved in LD formation and clustering in INS-1 β cells. More importantly, pre-treatment of INS-1 β cells with IFN-γ markedly increased PIC-induced expression of antiviral genes (e.g. Ifnb, Mx1)14
Although induction of LDs has been documented to occur mainly in macrophage models, following infection with bacteria, the ability of viral infection of cells to induce the same response remains relatively unexplored.
Recently, viral infection of the positive-stranded RNA viruses, Sindbis and dengue virus, was shown to induce LD formation in the cells of mosquito midgut for the first time15. This LD induction was mimicked via synthetic activation of the antiviral innate pathways, Toll and IMD, similar to the induction of bacterial-induced LDs.
Although it is known that activation of early innate signalling pathways appears to induce LDs in the presence of bacteria, and in the mosquito midgut when virally infected, the mechanisms at play remain unknown, as does the functional outcome of this LD induction. Here we show for the first time that LDs are induced early following both RNA and DNA viral infection and that this induction is transient in nature and facilitates an effective antiviral response.
References
- 1 Arrese, E. L., Saudale, F. Z. & Soulages, J. L. Lipid Droplets as Signaling Platforms Linking Metabolic and Cellular Functions. Lipid Insights 7, 7–16 (2014).PubMedGoogle Scholar
- 2 den Brok, M. H., Raaijmakers, T. K., Collado-Camps, E. & Adema, G. J. Lipid Droplets as Immune Modulators in Myeloid Cells. Trends Immunol. 39, 380–392 (2018).CrossRefGoogle Scholar
- 3 Gao, Q. & Goodman, J. M. The lipid droplet-a well-connected organelle. Front Cell Dev Biol 3, 49 (2015).Google Scholar
- 4 Henne, W. M., Reese, M. L. & Goodman, J. M. The assembly of lipid droplets and their roles in challenged cells. EMBO J. 38, (2019).Google Scholar
- 5 Mattos, K. A. et al. Modulation of lipid droplets by Mycobacterium leprae in Schwann cells: a putative mechanism for host lipid acquisition and bacterial survival in phagosomes. Cell. Microbiol. 13, 259–273 (2011).CrossRefPubMedGoogle Scholar
- 6 Rank, R. G., Whittimore, J., Bowlin, A. K. & Wyrick, P. B. In vivo ultrastructural analysis of the intimate relationship between polymorphonuclear leukocytes and the chlamydial developmental cycle. Infect. Immun. 79, 3291–3301 (2011).Abstract/FREE Full TextGoogle Scholar
- 7 Nicolaou, G., Goodall, A. H. & Erridge, C. Diverse bacteria promote macrophage foam cell formation via Toll-like receptor-dependent lipid body biosynthesis. J. Atheroscler. Thromb. 19, 137–148 (2012).CrossRefPubMedGoogle Scholar
- 8 D’Avila, H. et al. Host Cell Lipid Bodies Triggered by Trypanosoma cruzi Infection and Enhanced by the Uptake of Apoptotic Cells Are Associated With Prostaglandin E2 Generation and Increased Parasite Growth. The Journal of Infectious Diseases vol. 204 951–961 (2011).CrossRefPubMedGoogle Scholar
- 9 Tanigawa, K. et al. Expression of adipose differentiation-related protein (ADRP) and perilipin in macrophages infected with Mycobacterium leprae. FEMS Microbiol. Lett. 289, 72–79 (2008).CrossRefPubMedGoogle Scholar
- 10 Pacheco, P. et al. Lipopolysaccharide-induced leukocyte lipid body formation in vivo: innate immunity elicited intracellular Loci involved in eicosanoid metabolism. J. Immunol. 169, 6498–6506 (2002).Abstract/FREE Full TextGoogle Scholar
- 11 Dutta, A., Banerjee, S. & Sinha, D. K. Lipid Droplet metabolism dependent microbial defense in pre-immune zebrafish embryos. bioRxiv 478859 (2018) doi:10.1101/478859.Abstract/FREE Full TextGoogle Scholar
- 12 Anand, P. et al. A novel role for lipid droplets in the organismal antibacterial response. Elife 1, e00003 (2012).CrossRefGoogle Scholar
- 13 Knight, M., Braverman, J., Asfaha, K., Gronert, K. & Stanley, S. Lipid droplet formation in Mycobacterium tuberculosis infected macrophages requires IFN-γ/HIF-1α signaling and supports host defense. PLoS Pathog. 14, e1006874 (2018).CrossRefGoogle Scholar
- 14 Truong, N., Love-Rutledge, S., Lydic, T. & Olson, L. K. Effect of interferon gamma on neutral lipid levels, lipid droplet formation, and antiviral responses in pancreatic islets and INS-1 ß cells. The FASEB Journal 33, 654. 11–654. 11 (2019).Google Scholar
reference link : https://www.biorxiv.org/content/10.1101/2020.02.12.946749v1.full
More information: M. Bosch el al., “Mammalian lipid droplets are innate immune hubs integrating cell metabolism and host defense,” Science (2020). science.sciencemag.org/cgi/doi … 1126/science.aay8085



{kind=link}