











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A team of researchers from the Albert Einstein College of Medicine and the University of North Carolina at Chapel Hill has discovered that nociceptive nerves found in bone marrow help to regulate hematopoietic stem cell mobilization.
In their paper published in the journal Nature, the group describes their work, in which they attempted to learn more about nociceptive nerves found in bone marrow.
Anastasia Tikhonova and Iannis Aifantis with the University Health Network, Toronto and New York University’s Grossman School of Medicine, respectively, have published a News and Views piece on the work done by the researchers in the same journal issue.
As Tikhonova and Aifantis note, hematopoietic stem cells (HSCs) originate in bone marrow – they grow into blood and immune cells, which make their way into the bloodstream via a process called hematopoiesis. Prior to mobilization, HSCs reside in special niches inside of bone marrow – they only mobilize when needed.
In people being treated for leukemia and other blood cancers, HSCs are collected from the bone marrow using special drugs and stored for the duration of chemotherapy. Then the HSCs are replaced so that they can replenish the immune cells that were lost.
Unfortunately, in some people, collecting HSCs can be problematic because the HSCs refuse to be enticed out of their niches.
In this new effort, the researchers were studying nociceptive nerves in bone marrow hoping to learn why they exist in bone marrow – in other parts of the body, such as the skin, they play a role in pain sensation. The researchers found that they play a role in hematopoietic stem cell mobilization.
As part of their work, the researchers were using immunofluorescent imaging to study nerve cells in the bone marrow of mice, and discovered that most of them were nociceptive nerves. That led them to wonder if these nerves played a role in hematopoietic stem cell mobilization.
To find out if that might be the case, they used drugs and genetic engineering to disable them in two ways. They found that doing so had no impact on the maintenance of HSCs, but it did result in dramatically reduced hematopoietic stem cell mobilization.
The researchers then tried giving the mice drugs that increased production of the neurotransmitters associated with nociceptive nerves and found that doing so increased hematopoietic stem cell mobilization, as well. Intrigued, they also gave the mice capsaicin, a component in chili pepper, and found that it also increased hematopoietic stem cell mobilization.
Hematopoietic stem cells (HSC) reside in the bone marrow (BM) within a specialized micro-environment, the HSC niche, which comprises several cellular constituents. These include cells of mesenchymal origin, endothelial cells and HSC progeny, such as megakaryocytes and macrophages.
The BM niche and its cell populations ensure the functional preservation of HSCs. During infection or systemic inflammation, HSCs adapt to and respond directly to inflammatory stimuli, such as pathogen-derived signals and elicited cytokines, in a process termed emergency myelopoiesis, which includes HSC activation, expansion, and enhanced myeloid differentiation.
The cell populations of the niche participate in the regulation of emergency myelopoiesis, in part through secretion of paracrine factors in response to pro-inflammatory stimuli, thereby indirectly affecting HSC function.
Here, we review the crosstalk between HSCs and cell populations in the BM niche, specifically focusing on the adaptation of the HSC niche to inflammation and how this inflammatory adaptation may, in turn, regulate emergency myelopoiesis.
Introduction: HSCs and Inflammation
Hematopoietic stem cells (HSC) have self-renewal capacity and give rise to all mature blood cells, a process defined as hematopoiesis. The maintenance of the rare HSC population in the bone marrow (BM) and the preservation of their functional properties is supported by a highly specialized microenvironment inside the BM, the HSC niche (1, 2).
Besides extracellular matrix, the HSC niche (Figure 1) comprises different cell populations, including mesenchymal stromal cells (MSC), endothelial cells, osteolineage cells as well as progeny of HSCs, such as macrophages and megakaryocytes (1–3). The niche supports HSCs either via direct adhesive interactions (between HSCs and the cells or the extracellular matrix of the niche) or through the secretion of factors that act in a paracrine manner.
Such factors with paracrine effects on HSCs include C-X-C motif chemokine ligand (CXCL)-12, thrombopoietin, transforming growth factor (TGF)-β1 or stem cell factor (SCF; also called Kit-ligand) (1, 2). The fine regulation of HSCs by the BM niche microenvironment promotes maintenance of HSC quiescence, which is critical for preservation of their self-renewal potential (1).
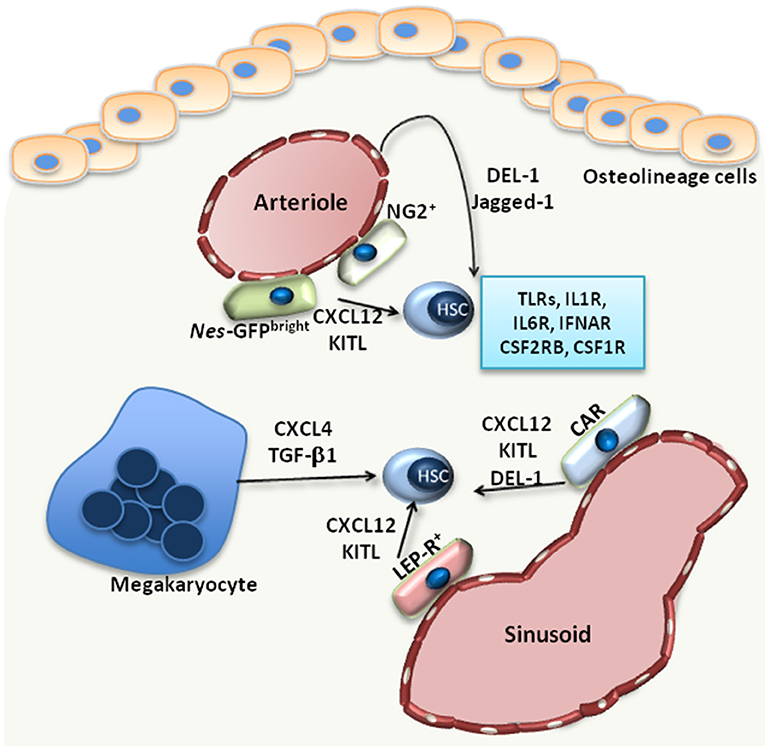
Hematopoietic stress drives HSCs to exit their quiescent state and undergo proliferation and lineage differentiation, according to the demands of the specific hematopoietic stress. In the case of systemic inflammation or infection, the resulting stress on hematopoiesis creates a tremendous need for production of mature myeloid cells, especially neutrophils and monocytes (3, 4).
This vital and pressing response of the BM to infections or peripheral inflammation is termed emergency myelopoiesis (5). Hematopoietic stem and progenitor cells (HSPCs) are fully equipped with pattern-recognition receptors (6), e.g., Toll-like receptors (TLR), and respond to pathogen-derived products that reach the BM through the circulation.
TLR activation of HSPCs leads to enhanced proliferation and myeloid cell production (6–8), thereby promoting mature myeloid cell replenishment in the course of infection (6). HSCs can also respond to several growth factors and cytokines released during inflammation by immune or other cells at inflamed sites or by cells within the BM microenvironment.
For instance, interleukin (IL)-1 or IL-6 act directly on HSCs, driving their proliferation and instructing their differentiation toward the myeloid lineage (9–11), while type I interferon [e.g., IFN-α induces the proliferation of HSCs (12). However, chronic exposure to cytokines may injure HSCs (9, 12, 13)].
Moreover, HSCs express the receptor for macrophage colony-stimulating factor (CSF1; also known as M-CSF), CSF1R, and respond to in vitro M-CSF stimulation with differentiation toward monocytes (14). The inflammatory adaptation of HSPCs can also contribute to induction of trained immunity, i.e., a non-specific memory of previous encounters that promotes enhanced responses by HSPCs and their progeny to future challenges (3, 15).
Specifically, agonists of trained immunity may stimulate the upregulation of the expression of the common β subunit of the granulocyte macrophage colony–stimulating factor/interleukin-3 receptor (CSF2RB), thereby promoting downstream signaling and leading to enhanced myelopoiesis (10). The direct effects of pathogen-derived factors, cytokines and growth factors on HSPCs have been recently reviewed elsewhere (3).
In the context of inflammatory stress, not only HSCs, but also cell populations that form the HSC niche, sense and respond to inflammatory stimuli, such as pathogen-derived products, cytokines or growth factors; this response is crucial for ensuring the steady replenishment of leukocytes (3–5). The focus of the present review is the adaptation of the HSC niche cells to inflammation. Specifically, we review here the role of HSC niche inflammation in emergency myelopoiesis and in the context of malignant hematopoiesis.
Conclusion
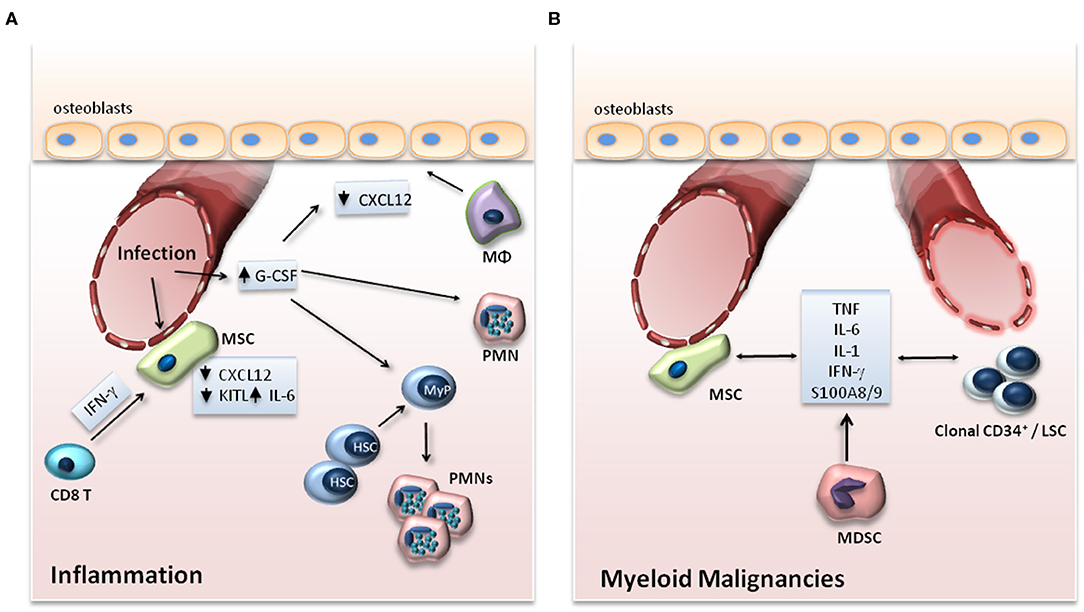
Under steady-state conditions, the crosstalk between cellular components of the niche and HSCs enables the maintenance of the latter in the BM in a state of quiescence, thereby ensuring the preservation of HSCs (2). During inflammation, HSCs are activated, proliferate, and preferentially differentiate toward the myeloid lineage (3). In parallel to the direct effect of inflammatory stimuli on HSCs, inflammatory alterations in niche cell populations and signals thereof also regulate the adaptation of HSCs to emergency myelopoiesis (3) (Figure 2A).
In addition, the interplay amongst HSCs, niche cell populations and inflammation is of critical importance in myeloid malignancies (54). The release of inflammatory mediators by clonal hematopoietic populations remodels the HSC niche in a manner that favors the preferential expansion of clonal leukemic cells, hence promoting the emergence and progression of malignant myeloid disease (54) (Figure 2B).
Therefore, it is imperative that future investigations focus on better understanding of how inflammation regulates the interactions of HSCs with their niche both during normal and malignancy-associated hematopoiesis.
reference link :https://www.frontiersin.org/articles/10.3389/fimmu.2020.01540/full
More information: Xin Gao et al. Nociceptive nerves regulate haematopoietic stem cell mobilization, Nature (2020). DOI: 10.1038/s41586-020-03057-y
Anastasia N. Tikhonova et al. Pain-sensing neurons mobilize blood stem cells from bone marrow, Nature (2020). DOI: 10.1038/d41586-020-03577-7



{kind=link}