










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



I risultati dello studio sono stati pubblicati sulla rivista peer review: Cellular And Molecular Immunology By Nature.
https://www.nature.com/articles/s41423-021-00779-5
I ricercatori hanno studiato l’ingresso delle cellule ospiti e la neutralizzazione mediata da anticorpi della variante A.30 (chiamata anche A.VOI.V2), che è stata rilevata in diversi pazienti in Angola e Svezia nella primavera del 2021 e probabilmente ha avuto origine in Tanzania [2]. Per confronto, abbiamo analizzato le varianti Beta (B.1.351) ed Eta (B.1.525).
Queste due varianti sono state rilevate per la prima volta in Africa e la variante Beta, che è considerata una variante preoccupante (VOC), mostra il più alto livello di resistenza alla neutralizzazione tra i VOC SARS-CoV-2 [3, 4] .
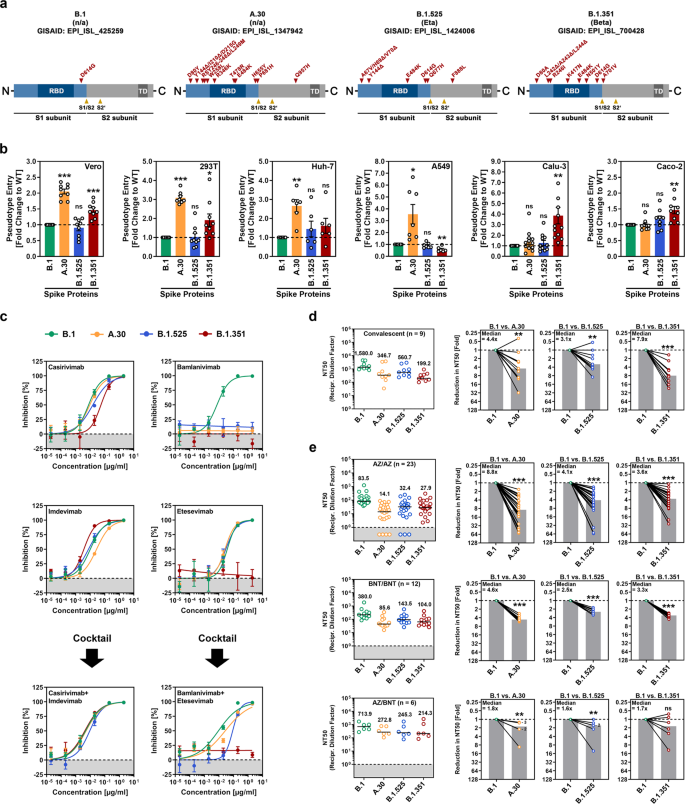
Rispetto alla proteina S di SARS-CoV-2 B.1, che circolava nella fase iniziale della pandemia, la proteina S della variante A.30 contiene 10 sostituzioni di amminoacidi e cinque delezioni (Fig. 1a e Informazioni supplementari, figura S1a).
Tutte le delezioni insieme a quattro sostituzioni si trovano nel dominio N-terminale dell’unità di superficie S1, che ospita un supersito antigenico che è preso di mira dalla maggior parte degli anticorpi neutralizzanti non diretti contro il dominio legante il recettore (RBD) [5].
Inoltre, tre mutazioni sono localizzate all’interno del RBD, che si lega al recettore cellulare ACE2 e costituisce il principale bersaglio degli anticorpi neutralizzanti (Fig. 1a).
Due di queste mutazioni, T478R ed E484K, si trovano vicino al sito di legame ACE2 (Informazioni supplementari, Fig. S1a) ed è noto che E484K riduce la suscettibilità alla neutralizzazione mediata da anticorpi. Infine, due mutazioni si trovano vicino al sito di scissione S1/S2 e una mutazione si trova nell’unità transmembrana S2, che facilita la fusione dell’involucro virale con le membrane cellulari (Fig. 1a).
Per l’analisi dell’ingresso virale nelle cellule e la sua inibizione da parte degli anticorpi, abbiamo impiegato pseudotipi rabdovirali recanti la proteina SARS-CoV-2 S, un modello adeguato per studiare l’ingresso e la neutralizzazione di SARS-CoV-2 [6].
Come bersagli, abbiamo usato le linee cellulari Vero e 293T (entrambe derivate dal rene), Huh-7 (fegato), A549 (polmone), Calu-3 (polmone) e Caco-2 (colon). B.1 è entrato in tutte le linee cellulari in modo efficiente e l’efficienza di ingresso di B.1.525 era comparabile (Fig. 1b e Informazioni supplementari, Fig. S1b).
L’ingresso di B.1.351 in diverse linee cellulari era leggermente ma significativamente aumentato e questo fenotipo era particolarmente robusto per le cellule polmonari Calu-3, in linea con i risultati pubblicati [3]. Infine, A.30 ha mostrato un’efficienza notevolmente aumentata per quanto riguarda l’ingresso nelle cellule Vero, 293 T, Huh-7 e A549 rispetto a B.1, sebbene l’ingresso nelle cellule Calu-3 e Caco-2 non sia stato aumentato (Fig. 1b).
Il test degli anticorpi monoclonali diretti contro la proteina S e utilizzati per la terapia COVID-19 ha rivelato che B.1.351 era resistente sia a bamlanivimab che a etesevimab, come previsto [3] e che B.1.525 era resistente a bamlanivimab (Fig. 1c). A.30 era anche resistente al bamlanivimab ma suscettibile di inibizione da parte di un cocktail di bamlanivimab ed etesevimab (Fig. 1c).
Inoltre, B.1.351 ha mostrato una neutralizzazione marcatamente ridotta da parte degli anticorpi indotti dall’infezione, come previsto; [3] L’evasione della neutralizzazione da parte di A.30 e B.1.525 era da leggermente (A.30) a moderatamente (B.1.525) meno efficiente (Fig. 1d e Informazioni supplementari, Fig. S2).
Al contrario, A.30 era più resistente alla neutralizzazione da parte degli anticorpi indotti sulla vaccinazione omologa ChAdOx1 nCoV-19 (Vaxzevria) o BNT162b2 (Comirnaty) rispetto a B.1.351, ma la sensibilità di neutralizzazione di B.1.525 era approssimativamente nello stesso intervallo di quella di B.1.351 (Fig. 1e e Informazioni supplementari, Fig. S2 e Tabella S1).
In sintesi, A.30 mostra una preferenza per la linea cellulare non osservata per altre varianti virali ed elude efficacemente la neutralizzazione da parte degli anticorpi provocati dalla vaccinazione ChAdOx1 nCoV-19 o BNT162b2.
L’ingresso di SARS-CoV-2 nelle linee cellulari dipende dall’attivazione della proteina S da parte delle proteasi cellulari catepsina L o TMPRSS2 [8] e si ritiene che l’attivazione da parte di quest’ultima supporti la diffusione virale nel polmone. Pertanto, è degno di nota che l’ingresso A.30 migliorato è stato osservato per le linee cellulari con catepsina L (cellule Vero, 293 T, Huh-7, A549), ma non l’ingresso dipendente da TMPRSS2 (Calu-3, Caco-2) [8 ].
Pertanto, si potrebbe ipotizzare che A.30 potrebbe utilizzare la catepsina L con maggiore efficienza e una leggera (ma non statisticamente significativa) resistenza di A.30 contro l’inibitore della catepsina L MDL 28170 supporta questa possibilità (Informazioni supplementari, Fig. S1c). In particolare, l’ingresso robusto nelle linee cellulari è stato combinato con un’elevata resistenza contro gli anticorpi indotti dalla vaccinazione con ChAdOx1 nCoV-19 o BNT162b2.
La resistenza alla neutralizzazione ha superato quella della variante Beta (B.1.351), che è marcatamente resistente alla neutralizzazione nella coltura cellulare e, rispetto alla variante Alpha (B.1.1.7), è meno ben inibita dal vaccino ChAdOx1 nCoV-19 [ 9]. Tuttavia, la vaccinazione eterologa ChAdOx1 nCoV-19/BNT162b2 , che in precedenza aveva dimostrato di aumentare le risposte anticorpali neutralizzanti contro i VOC rispetto alle corrispondenti vaccinazioni omologhe [7, 10], potrebbe offrire una solida protezione contro la variante A.30.
Collettivamente, i nostri risultati suggeriscono che la variante A.30 di SARS-CoV-2 può eludere il controllo da parte degli anticorpi indotti dal vaccino e potrebbe mostrare una maggiore capacità di entrare nelle cellule in modo dipendente dalla catepsina, il che potrebbe aiutare in particolare nella diffusione extrapolmonare. Di conseguenza, la potenziale diffusione della variante A.30 garantisce un attento monitoraggio e una rapida introduzione di contromisure.



{kind=link}