










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new study by researchers from the School of Clinical Medicine, Li Ka Shing Faculty of Medicine, the University of Hong Kong has found that SARS-CoV-2 infections also causes inflammatory bone loss leading to Long COVID skeletal health issues.
The study findings were published in the peer reviewed journal: Nature Communications.
https://www.nature.com/articles/s41467-022-30195-w
In addition to respiratory tract manifestations, extrapulmonary manifestations are also commonly reported in severe coronavirus infections such as COVID-19, SARS, and MERS4,26,27. Based on the phylogenetic similarities of SARS-CoV-2 and SARS-CoV, it has been postulated that the two beta coronaviruses may cause similar clinical features in infected patients.
The most severe musculoskeletal complication in SARS patients was non-progressive avascular necrosis of the femoral head caused by high-dose steroid pulse therapy 29.
During the revision of this work, it was also reported elsewhere that SARS-CoV-2 infection can induce bone loss in a lethal human ACE2-transgenic mouse model30,31,32. Nevertheless, it would be important to investigate the long-term bone changes not only in severe COVID-19 cases but also in mild to moderate ones, because most patients recover from the severe inflammation after proper treatment.
Thus, using our non-lethal hamster model represent mild to moderate human disease, we showed significant bone resorption happens at the acute inflammatory stage after SARS-CoV-2 infection. Moreover, in addition to a significantly increased number of RANK+ osteoclast precursors at this stage 33, there were also more TRAP+ osteoclasts expressing NFATc1, which is known to serve as a master regulator for terminal differentiation of osteoclasts 34.
This implies that pathological bone destruction may happen quickly after the onset of COVID-19. More importantly, similar findings were evident in different bone tissues harvested from the hamsters, suggesting that the bone loss is not site-specific but systemic. Without proper intervention, the bone volume and bone mineral density were barely restored even after the viral load became undetectable at the post-recovery/chronic inflammatory phase.
The presence of pathological bone loss may in turn complicate the rehabilitation of COVID-19 patients. For example, low bone mineral density is a known risk factor for vertebral fractures that may impair the respiratory function of COVID-19 patients in the rehabilitation phase 35,36. It was recently reported that thoracic vertebral fractures occurred in 36% of COVID-19 patients and increased the patients’ need for noninvasive mechanical ventilation 37.
Cytokine dysregulation has been associated with various clinical manifestations of COVID-19, including some involving the musculoskeletal system, such as myalgia, sarcopenia, arthralgia, and arthritis 39,40. Our findings in this study demonstrated that SARS-
CoV-2-induced pathological bone resorption through a pro-inflammatory cascade instead of direct infection in the skeletal tissue. Indeed, the direct infection of SARS-CoV-2 in bone tissue is very unlikely because there is little to no expression of ACE2 or co-expression of TMPRSS2, which are both vital for viral entry, in the bone marrow 41.
Additionally, a recent study demonstrated that SARS-CoV-2 was not able to bind and infect hematopoietic stem and progenitor cells in bone marrow, from which BMMs were derived 42. Consistent with our findings, it has been reported that while the viral load in the lung tissue of SARS-CoV-2-infected hamsters dropped from the peak level at 3 dpi to a significantly lower but still detectable level at 9 dpi, the viral load in bone marrow remained at the same level as mock control over the entire study period 43.
Multiplex quantification of immune mediators of SARS-CoV-2 peptide-stimulated cells from different organs also indicated that SARS-CoV-2–specific immune responses were primarily located in the lung or lung-associated lymph nodes instead of the bone marrow44.
After the inflammatory cytokines, (e.g., IL-1β and TNF-α) produced in the respiratory tract reached skeletal tissue via circulation, they quickly modulated the monocyte-macrophage lineage residing there to initiate additional pro-inflammatory cytokine cascade in the bone tissue.
The cytokines produced in the skeletal system, including but not limited to IFN-γ, IL-6, and PGE2, not only contributed to osteoclastogenesis in several interdependent signaling pathways 18 but also augmented the pro-osteoclastogenic actions of IL-1β and TNF-α by upregulating the expression of IL-1R1, which was implicated to primarily mediate pathological bone resorption 45.
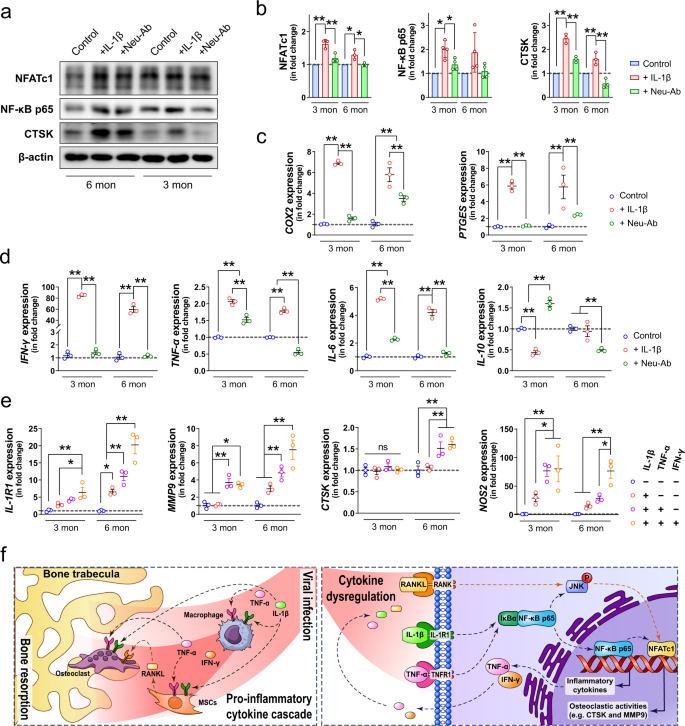
Among the various inflammatory factors that may be associated with osteoclastogenesis 22, we identified IL-1β and TNF-α as the key mediators for SARS-CoV-2-induced bone loss in the hamster model. As two of the most significantly upregulated inflammatory cytokines in the serum samples of SARS-CoV-2-infected hamsters, IL-1β and TNF-α have been known as potent stimulators of bone resorption from as early as the 1980s 46,47.
They are both shown to be essential in RANKL-induced osteoclast formation through the activation of osteoclastogenesis-related signaling pathways such as the c-Jun N-terminal Kinase (JNK) signaling 48,49. Besides their interdependent roles in mediating inflammatory osteopenia 50, IL-1β and TNF-α were also found to synergistically interact with other pro-inflammatory cytokines to stimulate osteoclastic differentiation 46,51,52.
In this study, we found that the concurrence of three pro-inflammatory cytokines (i.e., IL-1β, TNF-α, and IFN-γ) in the bone tissue after SARS-CoV-2 infection is preferably upregulated IL-1R1, which is primarily expressed in pathologically activated osteoclasts responsible for inflammatory bone destruction 53,54.
In contrast, the expression of IL-1R2, which serves as a decoy receptor for IL-1β to negatively regulate IL-1β signaling, was not significantly altered. Consequently, these SARS-CoV-2 infection-induced pro-inflammatory cytokines dramatically upregulated the expression of MMP9 and CTSK, which are both known to play dominant roles in the degradation of extracellular matrix 55.
Additionally, we also found that the inflammatory cytokines elevated in COVID-19 (e.g., IL‐1β) were able to promote the formation of osteoclasts via regulating the production of RANKL from MSCs, as reported elsewhere 56.
As we have demonstrated the SARS-CoV-2-induced bone loss in adult hamster, we further involved in vitro experiments using BMMs isolated from 3-month-old (young adult) or 6-month-old (matured adult) mice to test whether the inflammatory-mediated osteoclastogenesis is age-related.
Importantly, we demonstrated that BMMs isolated from young adult mice were more responsive to the pro-osteoclastic stimulation of IL‐1β. This is clinically relevant because young patients generally have a stronger ability to adequately respond to viral infections with rapid production of a high level of pro-inflammatory cytokines 57,58.
This heightened pro-inflammatory response, together with the lower baseline pro-inflammatory state in young patients, makes them more susceptible to various syndromes related to immune dysregulation58,59. Meanwhile, we also found that the involvement of other pro-inflammatory cytokines, such as TNF-α and IFN-γ, contributed to a more prominent effect on promoting the osteoclastic activities in BMMs isolated from matured adult mice than the ones from young mice.
This might explain why musculoskeletal symptoms are mostly seen in adult patients rather than children and the elderly60. Additionally, we noticed that the neutralizing antibody failed to block the late osteoclastic differentiation, especially in BMMs from young mice, as the size of osteoclasts and the expression of CTSK did not drop to the baseline level after the addition of the IL‐1β neutralizing antibody.
These data suggest that multiple neutralizing antibodies targeting different pro-inflammatory cytokines may need to be considered for limiting IL‐1β-induced osteoclastogenesis. As demonstrated in our study that the initial stimulation of IL‐1β led to an amplified and broadened pro-inflammatory cascade, resulting in the production of several other cytokines favoring osteoclastogenesis, simultaneous block of IL‐1β and other pro-inflammatory cytokines would be required for preventing bone loss61.
Besides the immunomodulatory effect and the pro-osteoclastogenesis effect, the accumulation of various pro-inflammatory cytokines in the skeletal tissue can lead to several other long-term health concerns.
For example, we showed that IL‐1β dramatically upregulated COX2 and PTGES, which both contribute to the production of PGE262. PGE2 is not only an inflammatory mediator involved in bone modeling but also a neuromodulator that can sensitize peripheral sensory neurons leading to inflammatory pain 63.
More than 20% of COVID-19 patients report lasting (from week 0 to week 28) bone ache or burning feeling10. Therefore, long-term monitoring of the inflammatory status of bone tissue after the recovery of the disease would be necessary. As a major indicator but not the sole driver in the pathology of COVID-1964, IL-6 is commonly found significantly increased in patients with exacerbating disease progression 64.
However, many studies have shown that the serum level of IL-6 would not be significantly upregulated in hamsters after SARS-CoV-2 infection 65,66, because SARS-CoV-2 would not cause severe or even lethal disease in hamsters. Instead, the virus would induce a mild to moderate pathologic procedure resembling the clinical situation found in most COVID-19 patients.
Therefore, our observation suggests that the inflammatory bone loss can happen in mild to moderate infection or post-recovery cases of COVID-19 patients. The inflammation in bone tissue can also alter the output of immune cells or cytokines from the bone marrow, which are supposed to participate in combating viral infection67. In our study, the significantly upregulated expression of IFN-γ and its signaling-associated genes (e.g., IRF1 and IRF2) in bone tissue indicate they might play an essential role in protecting bone tissue from virus infection 68.
However, it is also noteworthy that the expression of several anti-viral chemokines and cytokines, such as IFN-β69, IL-2170, and CCL1771, remained unchanged or even downregulated in the bone tissue. Thus, the suppression of the bone marrow-derived anti-viral factors may be one of the immune-evading mechanisms for SARS-CoV-2 that warrants further investigation.
In a recently published meta-analysis, Anakinra, a recombinant IL-1 receptor antagonist, was suggested as a safe, anti-inflammatory treatment option to reduce the mortality risk in patients admitted to hospital with moderate to severe COVID-19 pneumonia, especially in the presence of signs of hyperinflammation 72.
This suggests that adequate use of an anti-inflammatory treatment may be beneficial for COVID-19 during both the acute phase and the chronic phase involving long-term inflammatory complications. Therefore, evaluation of the effects of anti-inflammatory agents such as IL-1 receptor antagonists and IL-1 neutralizing antibodies on the prevention and/or treatment of SARS-CoV-2 infection-induced inflammatory bone loss should be included in future studies.
In addition, there are several limitations to our study. First. due to the lack of commercialized detection assays for evaluating biomolecules of hamster at the protein level, we only tested the circulating levels of IL-1β, TNF-α, and IL-6, which are known as three of the most important pro-inflammatory cytokines of the innate immune response.
Nevertheless, the striking similarities in the cytokine dysregulation between hamster model and human patients have been verified elsewhere using single-cell RNA sequencing73. Secondly, given the fact that the current hamster model mainly represents mild to moderate disease in human, additional investigations on the characteristics and treatment options for SARS-CoV-2-induced bone loss should be further conducted in more severe and/or lethal models.
In this study, we demonstrated the influence of SARS-CoV-2 infection on systemic bone loss during the acute and post-recovery/chronic phases. We revealed the pro-inflammatory cytokines derived from the respiratory system as the major mediators for pathological bone resorption. These pro-inflammatory cytokines disrupt the balance in bone metabolism and trigger another pro-inflammatory cascade in the skeletal tissue to further augment their pro-osteoclastogenesis effect (Fig. 8e).
The findings in our study highlight the need to closely monitor COVID-19 patients’ bone density. The benefits of prophylactic or therapeutic interventions against the development of pathological bone loss in COVID-19 patients should be further evaluated in animal models and clinical trials.



{kind=link}