










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Researchers at Utrecht University, Erasmus Medical Center and Harbour BioMed (HBM) today reported that they have identified a fully human monoclonal antibody that prevents the SARS-CoV-2 (COVID-19) virus from infecting cultured cells.
The discovery, published online today in Nature Communications, is an initial step towards developing a fully human antibody to treat or prevent the respiratory disease COVID-19 caused by the novel coronavirus SARS-CoV-2.
The COVID-19 pandemic has spread rapidly around the globe, infecting more than 3.3M people worldwide and killing more than 235,000 people so far.
“This research builds on the work our groups have done in the past on antibodies targeting the SARS-CoV that emerged in 2002/2003,” said Berend-Jan Bosch, Associate Professor, Research leader at Utrecht University, and co-lead author of the Nature Communications study.
“Using this collection of SARS-CoV antibodies, we identified an antibody that also neutralizes infection of SARS-CoV-2 in cultured cells.
Such a neutralizing antibody has potential to alter the course of infection in the infected host, support virus clearance or protect an uninfected individual that is exposed to the virus.”
Dr. Bosch noted that the antibody binds to a domain that is conserved in both SARS-CoV and SARS-CoV-2, explaining its ability to neutralize both viruses. “This cross-neutralizing feature of the antibody is very interesting and suggests it may have potential in mitigation of diseases caused by future-emerging related coronaviruses.”
“This discovery provides a strong foundation for additional research to characterize this antibody and begin development as a potential COVID-19 treatment,” said Frank Grosveld, Ph.D. co-lead author on the study, Academy Professor of Cell Biology, Erasmus Medical Center, Rotterdam and Founding Chief Scientific Officer at Harbour BioMed.
“The antibody used in this work is ‘fully human,” allowing development to proceed more rapidly and reducing the potential for immune-related side effects.” Conventional therapeutic antibodies are first developed in other species and then must undergo additional work to ‘humanize’ them.
The antibody was generated using Harbour BioMed’s H2L2 transgenic mouse technology.
“This is groundbreaking research,” said Dr. Jingsong Wang, Founder, Chairman & Chief Executive Officer of HBM. “Much more work is needed to assess whether this antibody can protect or reduce the severity of disease in humans.
We expect to advance development of the antibody with partners. We believe our technology can contribute to addressing this most urgent public health need and we are pursuing several other research avenues.”
Identification of SARS-CoV-2 reactive antibodies
In order to identify SARS-CoV-2-neutralizing antibodies, ELISA-(cross)reactivity was assessed of antibody-containing supernatants of a collection of 51 SARS-S hybridoma’s derived from immunized transgenic H2L2 mice that encode chimeric immunoglobulins with human variable heavy and light chains and constant regions of rat origin (Supplementary Table 1).
Four of 51 SARS-S hybridoma supernatants displayed ELISA-cross-reactivity with the SARS2-S1 subunit (S residues 1–681; Supplementary Table 1), of which one (47D11) exhibited cross-neutralizing activity of SARS-S and SARS2-S pseudotyped VSV infection.
The chimeric 47D11 H2L2 antibody was reformatted to a fully human immunoglobulin, by cloning of the human variable heavy and light chain regions into a human IgG1 isotype backbone. The recombinantly expressed human 47D11 was used for further characterization.
Antiviral and biochemical properties of the human mAb 47D11
The human 47D11 antibody binds to cells expressing the full-length spike proteins of SARS-CoV and SARS-CoV-2 (Fig. 1a). The 47D11 antibody was found to potently inhibit infection of VeroE6 cells with SARS-S and SARS2-S pseudotyped VSV with IC50 values of 0.061 and 0.061 μg/ml (Fig. 1b), respectively.
Authentic infection of VeroE6 cells with SARS-CoV and SARS-CoV-2 was neutralized with IC50 values of 0.19 and 0.57 μg/ml (Fig. 1c). Using ELISA 47D11 was shown to target the S1B receptor-binding domain (RBD) of SARS-S and SARS2-S. 47D11 bound the S1B of both viruses with similar affinities as shown by the ELISA-based half maximal effective concentration (EC50) values (0.02 and 0.03 μg/ml, respectively; Fig. 2a).
ELISA-based binding affinity of 47D11 for the spike ectodomain (Secto) of SARS-CoV was higher relative to that of SARS-CoV-2 (EC50 values: 0.018 and 0.15 μg/ml, respectively), despite equimolar antigen coating (Supplementary Fig. 1).
Congruent with the ELISA-reactivities, measurement of binding kinetics of 47D11 by biolayer interferometry showed that 47D11 binds SARS-Secto with higher affinity (equilibrium dissociation constant [KD]: 0.745 nM) relative to SARS2-Secto (KD 10.8 nM), whereas affinity for SARS-S1B and SARS2-S1B was in a similar range (16.1 and 9.6 nM, respectively, Supplementary Fig. 2).
This difference may originate from differences in epitope accessibility in SARS-S versus SARS2-S, as domain B can adopt a closed and open conformation in the prefusion spike homotrimer12,13.
Remarkably, binding of 47D11 to SARS-S1B and SARS2-S1B did not compete with S1B binding to the ACE2 receptor expressed at the cell surface as shown by flow cytometry (Fig. 2b; Supplementary Fig. 3) nor with Secto and S1B binding to soluble ACE2 in solid-phase based assay (Supplementary Fig. 4), whereas two SARS-S1 specific antibodies 35F4 and 43C6 that neutralize SARS-S (but not SARS2-S) pseudotyped VSV infection (Supplementary Fig. 5) do block binding of SARS-Secto and SARS-S1B to ACE2.
Using a trypsin-triggered cell-cell fusion assay, 47D11 was shown to impair SARS-S and SARS2-S mediated syncytia formation (Supplementary Fig. 6). Our data show that 47D11 neutralizes SARS-CoV and SARS-CoV-2 through a yet unknown mechanism that is different from receptor-binding interference.
Alternative mechanisms of coronavirus neutralization by RBD-targeting antibodies have been reported including spike inactivation through antibody-induced destabilization of its prefusion structure17, which may also apply for 47D11.
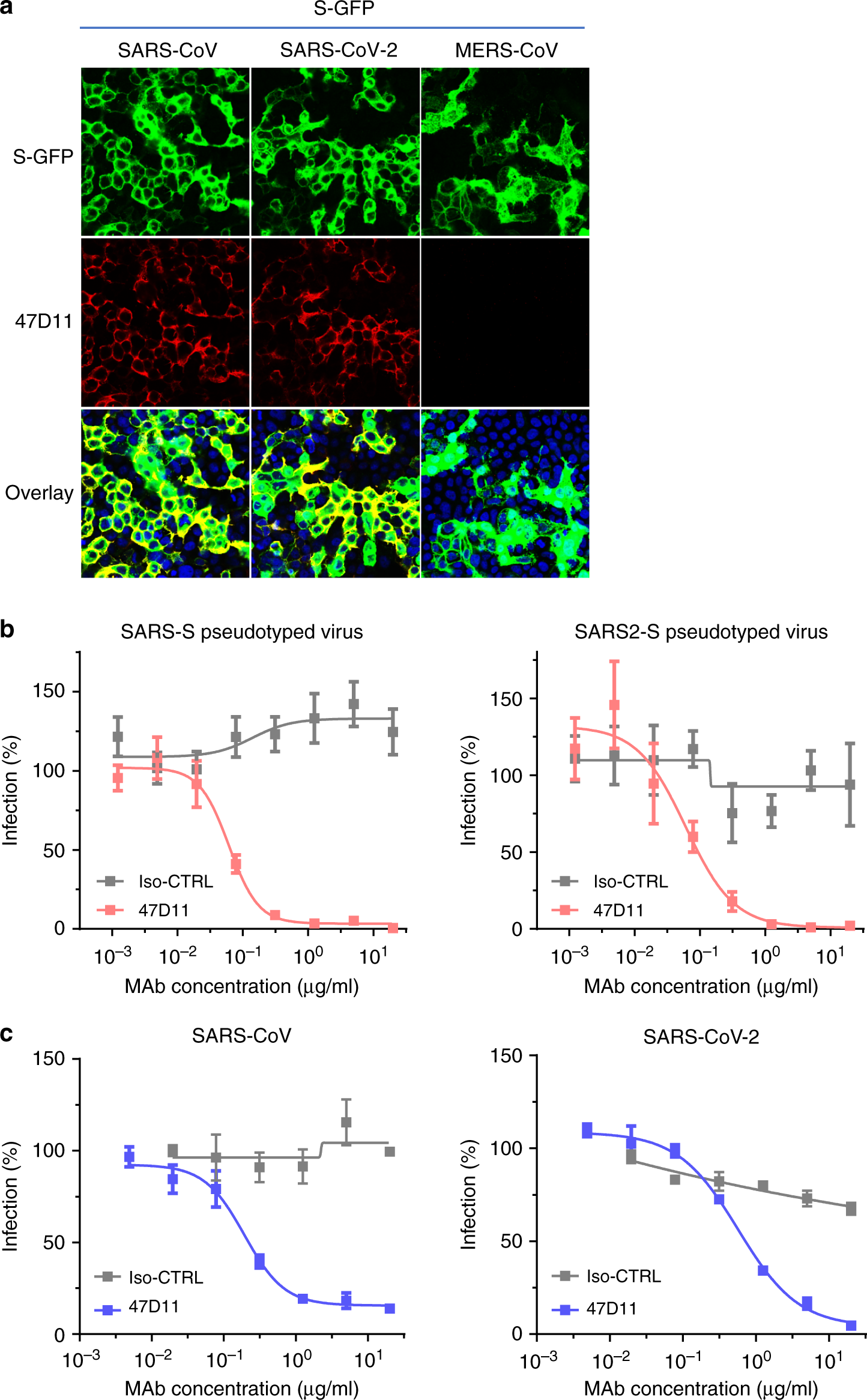
b Antibody-mediated neutralization of infection of luciferase-encoding VSV particles pseudotyped with spike proteins of SARS-CoV and SARS-CoV-2. Pseudotyped VSV particles pre-incubated with antibodies at indicated concentrations (see Methods) were used to infect VeroE6 cells and luciferase activities in cell lysates were determined at 24 h post transduction to calculate infection (%) relative to non-antibody-treated controls. The average ± SD from at least three independent experiments with technical triplicates is shown. Iso-CTRL: an anti-Strep-tag human monoclonal antibody11 was used as an antibody isotype control.
c Antibody-mediated neutralization of SARS-CoV and SARS-CoV-2 infection on VeroE6 cells. The experiment was performed with triplicate samples, the average ± SD is shown. Source data are provided as a Source Data file.
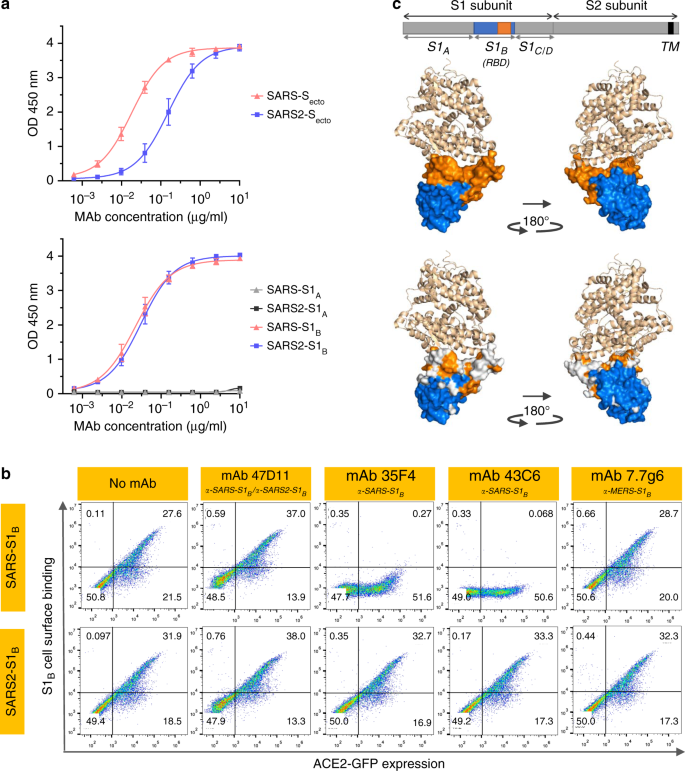
a ELISA-binding curves of 47D11 to Secto (upper panel) or S1A and S1B (RBD: receptor-binding domain) (lower panel) of SARS-S and SARS2-S coated at equimolar concentrations. The average ± SD from two independent experiments with technical duplicates is shown.
b Interference of antibodies with binding of the S-S1B of SARS-CoV and SARS-CoV-2 to cell surface ACE2-GFP analyzed by flow cytometry. Prior to cell binding, S1B was mixed with mAb (mAbs 47D11, 35F4, 43C6, 7.7G6, in H2L2 format) with indicated specificity in a mAb:S1B molar ratio of 8:1 (see Supplementary Fig. 3 for an extensive analysis using different mAb:S1B molar ratio’s). Cells are analyzed for (ACE2-)GFP expression (x axis) and S1B binding (y axis). Percentages of cells that scored negative, single positive, or double positive are shown in each quadrant. Experiment was done twice, a representative experiment is shown.
c Divergence in surface residues in S1B of SARS-CoV and SARS-CoV-2. Upper panel: Structure of the SARS-CoV spike protein S1B RBD in complex with human ACE2 receptor (PDB: 2AJF)24. ACE2 (wheat color) is visualized in ribbon presentation. The S1B core domain (blue) and subdomain (orange) are displayed in surface presentation using PyMOL, and are visualized with the same colors in the linear diagram of the spike protein above, with positions of the S1 and S2 subunits, the S ectodomain (Secto), the S1 domains S1A-D and the transmembrane domain (TM) indicated. Lower panel: similar as panel above with surface residues on S1B of SARS-CoV that are at variance with SARS-CoV-2 colorored in white. Source data are provided as a Source Data file.
More information: A human monoclonal antibody blocking SARS-CoV-2 infection, Nature Communications (2020). DOI: 10.1038/s41467-020-16256-y , www.nature.com/articles/s41467-020-16256-y



{kind=link}