









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


Yunjeong Kim e Kyeong-Ok “KC” Chang, virologi del College of Veterinary Medicine della Kansas State University, hanno pubblicato uno studio che mostra un possibile trattamento terapeutico per COVID-19 .
I coronavirus patogeni sono una grave minaccia per la salute pubblica globale, come dimostrato dalla sindrome respiratoria acuta grave coronavirus o SARS-CoV ; Sindrome respiratoria del Medio Oriente coronavirus, nota come MERS-CoV; e il SARS-CoV-2 appena emerso, il virus che causa l’infezione da COVID-19.
Lo studio, “Gli inibitori della proteasi simili al 3C bloccano la replicazione del coronavirus in vitro e migliorano la sopravvivenza nei topi con infezione da MERS-CoV “, appare nel numero del 3 agosto della prestigiosa rivista medica Science Translational Medicine .
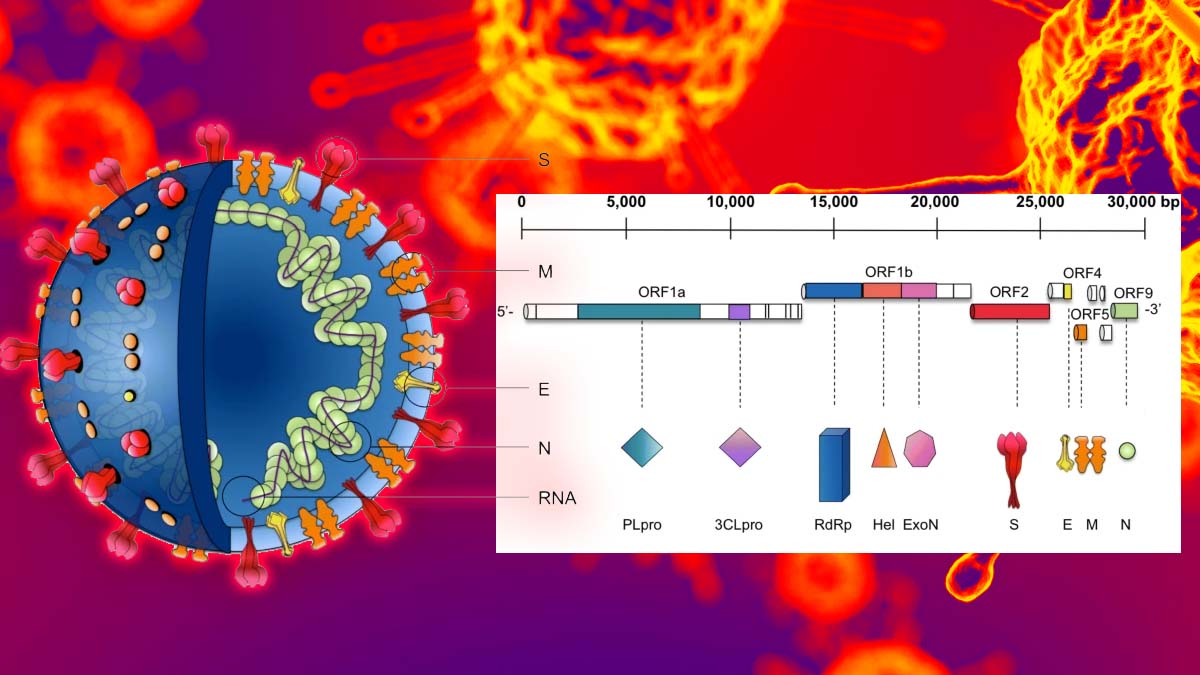
Rivela come gli inibitori della proteasi delle piccole molecole mostrano la potenza contro i coronavirus umani. Queste proteasi simili al 3C di coronavirus, note come 3CLpro, sono forti bersagli terapeutici perché svolgono ruoli vitali nella replicazione del coronavirus.
“Gli sviluppi e i trattamenti dei vaccini sono i principali obiettivi della ricerca COVID-19 e il trattamento è davvero fondamentale”, ha affermato Chang, professore di medicina diagnostica e patobiologia.
“Questo documento descrive gli inibitori della proteasi che colpiscono il coronavirus 3CLpro, che è un noto bersaglio terapeutico.”
Lo studio dimostra che questa serie di inibitori ottimizzati del coronavirus 3CLpro hanno bloccato la replicazione dei coronavirus umani MERS-CoV e SARS-CoV-2 nelle cellule in coltura e in un modello murino per MERS.
Questi risultati suggeriscono che questa serie di composti dovrebbe essere ulteriormente studiata come potenziale terapeutico per l’infezione da coronavirus umano.
Chang e Kim hanno utilizzato le sovvenzioni del National Institutes of Health per sviluppare farmaci antivirali per il trattamento di MERS e infezioni da norovirus umano. Il loro lavoro si estende ad altri virus umani come i rinovirus e la SARS-CoV-2.
“Il lavoro che questo gruppo di collaboratori ha svolto su antivirali e inibitori per SARS e MERS presso K-State da diversi anni è stato vitale per la loro capacità di ruotare rapidamente per enfatizzare la ricerca sul virus e sulle terapie SARS-CoV-2, “Ha dichiarato Peter K. Dorhout, vicepresidente per la ricerca presso il K-State.
Collaboratori della ricerca includono team guidati da Bill Groutas alla Wichita State University, Stanley Perlman all’Università dello Iowa e Scott Lovell all’Università del Kansas.
“Drs. Groutas, Perlman e Lovell hanno portato decenni di esperienza nel nostro team di ricerca “, ha affermato Chang. “Non saremmo stati in grado di arrivare così lontano senza importanti collaborazioni con i nostri colleghi di altre istituzioni”.
“Pubblicare le cose in questo momento è molto importante per la comunità scientifica”, ha detto Kim. “Penso che stiamo aggiungendo preziose informazioni al campo antivirale.”
Lo scoppio del coronavirus SARS-CoV-2 a Wuhan, in Cina, nel dicembre 2019, la causa della malattia da Corona Virus del 2019 (COVID-19), rappresenta una minaccia pandemica per la salute globale [1,2]. L’OMS ha dichiarato COVID-19 come una pandemia l’11 marzo 2020.
L’epidemia si è diffusa in oltre 185 paesi con oltre 3.200.000 casi confermati, oltre 230.000 decessi confermati e oltre 1.000.000 di recuperi totali in tutto il mondo dal 1 ° maggio 2020 [3].
Centinaia di milioni di vite sono state colpite a causa di isolamenti / quarantene obbligatori. Questa pandemia ha il potenziale per sopraffare i sistemi sanitari nazionali e avere conseguenze importanti sull’economia globale se la diffusione e la virulenza della SARS-CoV-2 non sono contenute o se non vengono sviluppati trattamenti efficaci. I coronavirus sono raggruppati in alfa, beta, gamma e delta classi. I coronavirus possono infettare sia l’uomo che gli animali.
Si ritiene che la fonte del beta coronavirus SARS-CoV-2 siano i pipistrelli, che trasportano il virus senza segni di malattia [4]. I beta-coronavirus hanno causato epidemie precoci di sindromi respiratorie acute acute (SARS), tra cui SARS-CoV (2002/2003 nel Guangdong, Cina) e il virus della sindrome respiratoria mediorientale MERS-CoV (2012 in Arabia Saudita) [5].
I beta-coronavirus sono patogeni per l’uomo e hanno un genoma a RNA a singolo filamento, incapsulato da un involucro di membrana [6]. La morfologia a forma di corona di coronavirus (“corona”) è creata da glicoproteine a spiga transmembrana (proteine S) che formano omotrimeri che sporgono dalla superficie virale [7].
Le proteine S di SARS-CoV e SARS-CoV-2 mostrano l’omologia strutturale e gli ectodomini conservati, quindi strategie precedenti impiegate per prevenire il legame di SARS-CoV al suo recettore della cellula ospite dell’enzima di conversione dell’angiotensina 2 (ACE2) possono essere rilevanti, poiché SARS -CoV-2 impiega anche ACE2 per l’immissione di celle [8,9].
L’ACE2, una esopeptidasi espressa sulle cellule epiteliali del tratto respiratorio, può costituire un obiettivo farmacologico per limitare l’ingresso cellulare di SARS-CoV-2. È stato dimostrato che i farmaci antimalarici consolidati clorochina e idrossiclorochina inibiscono la fosforilazione terminale di ACE2 e aumentano il pH negli endosomi, rispettivamente.
La clorochina e l’idrossiclorochina costituiscono farmaci candidati contro l’infezione da SARS-CoV-2 e la malattia COVID-19, e sono ora studiati per la loro efficacia terapeutica negli studi clinici internazionali con pazienti COVID-19 (ovvero la prova SOLIDARITY).
La proteina S glicosilata di SARS-CoV-2 è altamente immunogenica per il sistema immunitario ospite e gli anticorpi policlonali murini contro la proteina SARS-Co-V S inibiscono potentemente l’ingresso di cellule mediate da SARS-CoV-2 S, indicando che gli anticorpi neutralizzanti l’incrocio prendendo di mira gli epitopi S conservati possono essere sollecitati dopo la vaccinazione [9].
Simile ai precedenti beta-coronavirus SARS e MERS, la SARS-CoV-2 infetta principalmente le cellule epiteliali alveolari del polmone, portando a una polmonite periferica bilaterale grave con opacità del vetro smerigliato nelle immagini CT (malattia COVID-19), con un tasso di mortalità di Dal 2% al 5% [10].
Anche la SARS-CoV-2 può contribuire all’insufficienza multipla di organi, interessando cuore, fegato, reni, sistema nervoso centrale e tratto gastrointestinale [11]. L’epidemiologia finora suggerisce che SARS-CoV-2 è più contagioso di SARS-CoV o MERS-CoV [12].
Numerosi meccanismi ora identificati nei processi infettivi e di replicazione di SARS-CoV-2 offrono obiettivi per interventi farmacologici. L’infezione di pneumociti, macrofagi e mastociti polmonari richiede la proteina S virale.
Questo processo di invasione che prevede l’attacco della proteina S al recettore ACE2 è facilitato dalla proteasi serinica derivata dalle cellule ospiti TMPRSS2 [8]. Gli agenti che inibiscono TMPRSS2, come il camilato mesilato, possono essere utili nel bloccare l’ingresso della cellula ospite virale.
Dopo l’ingresso della cellula ospite, l’RNA positivo a singolo filamento virale viene rilasciato per la replicazione dell’RNA virale e la traduzione delle polipoteine virali che vengono infine suddivise in proteine effettrici mature dalle proteasi virali [13].
L’interazione della proteina S con ACE2 sulla membrana citoplasmatica della cellula ospite avvia l’infezione virale. Le strategie in grado di interrompere l’interazione della proteina S con ACE2 potrebbero avere un significativo valore terapeutico, poiché l’affinità di legame della proteina S SARS-CoV-2 con ACE2 è 10-20 volte superiore rispetto alla proteina S di SARS-CoV che può contribuire a la maggiore contagiosità di SARS-CoV-2 rispetto a SARS-CoV [12].
Sebbene SARS-CoV e SARS-CoV-2 abbiano solo il 79% di somiglianza nella sequenza genomica, condividono un dominio di legame recettoriale altamente conservato per le loro proteine S [1].
Esiste anche il potenziale per colpire altre proteine altamente conservate associate a SARS-CoV e SARS-Co-V-2, tra cui RdRp e 3Clpro (anche chiamato Mpro), che condividono oltre il 95% di somiglianza tra i due virus, nonostante solo il 79% di genomica condivisione sequenziale.
RdRp è una RNA polimerasi RNA-dipendente richiesta per replicare il genoma virale all’interno della cellula ospite. 3Clpro e Plpro sono entrambe proteasi virali che scompongono la polipoteina virale in unità funzionali all’interno delle cellule ospiti che vengono infine assemblate in nuovi virus.
Le sequenze 3Clpro tra i due virus sono simili al 96%, l’identità della sequenza Plpro è dell’83% e i loro siti attivi mostrano un alto grado di conservazione [14].
I farmaci che hanno recentemente dimostrato di colpire MERS-CoV nei topi [15] e di inibire il virus Ebola RdRp e le proteasi SARS-CoV-2 nell’uomo, come remdesivir e ritonavir / lopinavir, costituiscono anche farmaci candidati contro SARS-CoV- 2 e sono ora studiati per la loro efficacia terapeutica nei pazienti COVID-19 in 2 studi clinici internazionali (SOLIDARITY Trial e DisCoVeRy Trial).
Infine, per il trattamento dell’infezione da SARS-CoV-2 e della malattia COVID-19 dovrebbero essere presi in considerazione alcuni prodotti fitochimici e prodotti naturali ad alta attività antivirale.

Inibitori della proteasi SARS-CoV-2 3Clpro
3Clpro (anche chiamato Mpro) costituisce la principale proteasi dei beta coronavirus che è essenziale per l’elaborazione delle polipoteine tradotte dall’RNA virale [125]. Recentemente, sono state segnalate le strutture a raggi X del 3Clpro SARS-CoV-2 non obbligato e il suo complesso con α-chetoamidi progettati come inibitori specifici di 3Clpro [126].
Due α-chetoamidi contenenti piridina, designate 13a e 13b, hanno mostrato proprietà farmacocinetiche favorevoli nei topi e sono state rilevate a concentrazioni sufficienti nel tessuto polmonare e nel fluido di lavaggio broncheo-alveolare entro 4 ore-24 ore dopo la somministrazione sottocutanea, dimostrando tropismo polmonare dei composti [126].
Oltre alla somministrazione sottocutanea, l’inalazione di 13b nebulizzato da parte dei topi ha comportato concentrazioni elevate e di lunga durata (24 ore) nel tessuto polmonare, senza effetti avversi [126], sottolineando un ruolo delle α-chetoamidi contenenti piridina nella terapia COVID-19 .
In uno studio recente che ha utilizzato la progettazione combinata di farmaci assistiti da struttura, screening di droghe virtuali e screening ad alto rendimento, un inibitore basato sul meccanismo di 3Clpro, chiamato N3, è stato identificato dal design computerizzato di farmaci [127].
N3, un inibitore dell’accettore di Michael che può inibire i 3Clpros di SARS-CoV e MERS-CoV, ha dimostrato di formare un legame covalente con e di essere un inibitore irreversibile di SARS-CoV-2 3Clpro [127].
Inoltre, in un approccio di screening ad alto rendimento per l’identificazione di inibitori della SARS-CoV-2 3Clpro, è stato identificato ebselen, un composto di organoselenium con proprietà antinfiammatorie, antiossidanti e citoprotettive [127].
In un saggio di riduzione della placca con cellule Vero simiane infettate con SARS-CoV-2, N3 ed ebselen hanno mostrato un’efficacia antivirale e di protezione cellulare a valori EC50 di 16,77 μM e 4,67 μM, rispettivamente [127], dimostrando infine il loro potenziale antivirale contro SARS- CoV-2.
Riferimenti
1. Lu R., Zhao X., Li J., Niu P., Wang B., Wu H. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet. 2020;395(10224):565–574. [PMC free article] [PubMed] [Google Scholar]
2. Zhu N., Zhang D., Wang W., Li X., Yang B., Song J. A novel coronavirus from patients with pneumonia in China. N. Engl. J. Med. 2019;383(8):727–733. 2020. [Google Scholar]
3. https://coronavirus.jhu.edu/map.html Johns Hopkins University, USA, (2020).4. Li C., Yang Y., Ren L. Genetic evolution analysis of 2019 novel coronavirus and coronavirus from other species. Infect. Genet. Evol. 2020;82:104285. [PMC free article] [PubMed] [Google Scholar]
5. Anthony S.J., Johnson C.K., Greig D.J., Kramer S., Che X., Wells H. Global patterns in coronavirus diversity. Virus Evol. 2017;3(1):vex012. [PMC free article] [PubMed] [Google Scholar]
6. Su S., Wong G., Shi W., Liu J., Lai A.C.K., Zhou J. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016;24(6):490–502. [PMC free article] [PubMed] [Google Scholar]
7. Tortorici M.A., Veesler D. Structural insights into coronavirus entry. Adv. Virus Res. 2019;105:93–116. [PMC free article] [PubMed] [Google Scholar]
8. Hoffmann M., Kleine-Weber H., Schroeder S., Krüger N., Herrler T., Erichsen S. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease receptor. Cell. 2020;(March 4) doi: 10.1016/j.cell.2020.02.052. pii: S0092-8674(20)30229-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Walls A.C., Park Y.J., Tortorici M.A., Wall A., McGuire A.T., Veesler D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. 2020;(March 6) doi: 10.1016/j.cell.2020.02.058. pii: S0092-8674(20)30262-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
10. Wu Y.C., Chen C.S., Chan Y.J. The outbreak of COVID-19: an overview, J. Chin. Med. Assoc. 2020;83(2):217–220. [PMC free article] [PubMed] [Google Scholar]
11. Zhu N., Zhang D., Wang W., Li X., Yang B., Song J. A novel coronavirus from patients with pneumonia in China. N. Engl. J. Med. 2020;382(8):727–733. [PMC free article] [PubMed] [Google Scholar]
12. Tang B., Bragazzi N.L., Li Q., Tang S., Xiao Y., Wu J. An updated estimation of the risk of transmission of the novel coronavirus (2019-nCov) Infect. Dis. Model. 2020;5:248–255. [PMC free article] [PubMed] [Google Scholar]
13. Báez-Santos Y.M., St John S.E., Mesecar A.D. The SARS-coronavirus papain-like protease: structure, function and inhibition by designed antiviral compounds. Antiviral Res. 2015;115:21–38. [PMC free article] [PubMed] [Google Scholar]
14. Morse J.S., Lalonde T., Xu S., Liu W.R. Learning from the past: possible urgent prevention and treatment options for severe acute respiratory infections caused by 2019-nCoV. Chembiochem. 2020;21(5):730–738. [PMC free article] [PubMed] [Google Scholar]
124. NCT04303299, ClinicalTrials.gov, (2020), Mar 11.
125. Anand K., Ziebuhr J., Wadhwani P., Mesters J.R., Hilgenfeld R. Coronavirus main protease (3CLpro) structure: basis for design of anti-SARS drugs. Science. 2003;300(5626):1763–1767. [PubMed] [Google Scholar]
126. Zhang L., Lin D., Sun X., Curth U., Drosten C., Sauerhering L. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science. 2020;(March 20) doi: 10.1126/science.abb3405. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
127. Jin Z., Du X., Xu Y., Deng Y., Liu M., Zhao Y. Structure of Mpro from COVID-19 virus and discovery of its inhibitors. Nature. 2020;(April 9) doi: 10.1038/s41586-020-2223-y. [PubMed] [CrossRef] [Google Scholar]
More information: “3C-like protease inhibitors block coronavirus replication in vitro and improve survival in MERS-CoV-infected mice,” Science Translational Medicine (2020). DOI: 10.1126/scitranslmed.abc5332 , stm.sciencemag.org/content/ear … scitranslmed.abc5332



{kind=link}