










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Humans are not the only species facing a potential threat from SARS-CoV-2, the novel coronavirus that causes COVID-19, according to a new study from the University of California, Davis.
An international team of scientists used genomic analysis to compare the main cellular receptor for the virus in humans – angiotensin converting enzyme-2, or ACE2 – in 410 different species of vertebrates, including birds, fish, amphibians, reptiles and mammals.
ACE2 is normally found on many different types of cells and tissues, including epithelial cells in the nose, mouth and lungs.
In humans, 25 amino acids of the ACE2 protein are important for the virus to bind and gain entry into cells.
The researchers used these 25 amino acid sequences of the ACE2 protein, and modeling of its predicted protein structure together with the SARS-CoV-2 spike protein, to evaluate how many of these amino acids are found in the ACE2 protein of the different species.
“Animals with all 25 amino acid residues matching the human protein are predicted to be at the highest risk for contracting SARS-CoV-2 via ACE2,” said Joana Damas, first author for the paper and a postdoctoral research associate at UC Davis. “The risk is predicted to decrease the more the species’ ACE2 binding residues differ from humans.”
About 40 percent of the species potentially susceptible to SARS-CoV-2 are classified as “threatened” by the International Union for Conservation of Nature and may be especially vulnerable to human-to-animal transmission. The study was published Aug. 21 in the Proceedings of the National Academy of Sciences.
“The data provide an important starting point for identifying vulnerable and threatened animal populations at risk of SARS-CoV-2 infection,” said Harris Lewin, lead author for the study and a distinguished professor of evolution and ecology at UC Davis. “We hope it inspires practices that protect both animal and human health during the pandemic.”
Endangered species predicted to be at risk
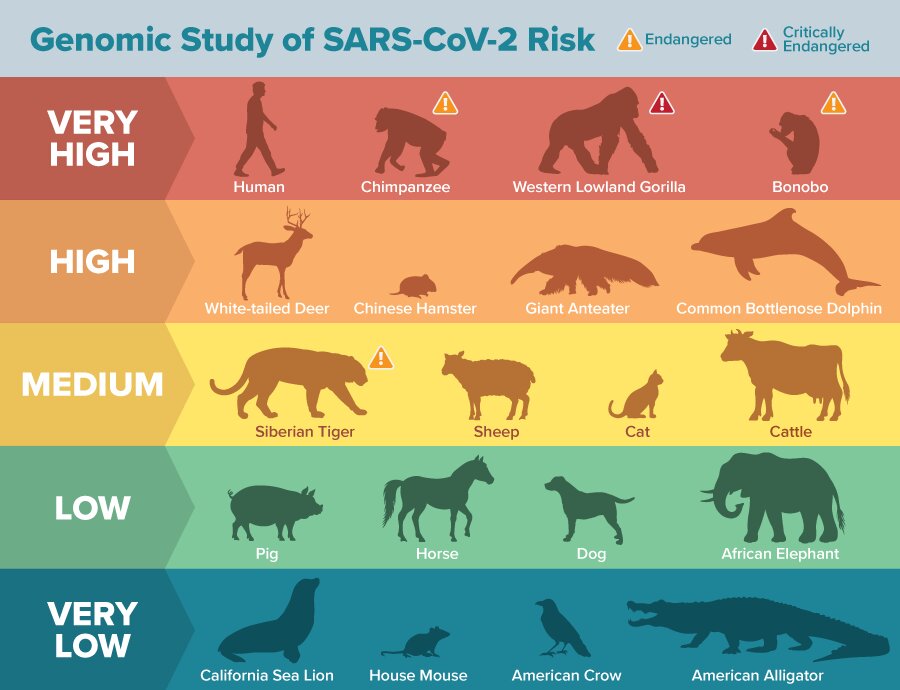
Several critically endangered primate species, such as the Western lowland gorilla, Sumatran orangutan and Northern white-cheeked gibbon, are predicted to be at very high risk of infection by SARS-CoV-2 via their ACE2 receptor.
Other animals flagged as high risk include marine mammals such as gray whales and bottlenose dolphins, as well as Chinese hamsters.
Domestic animals such as cats, cattle and sheep were found to have a medium risk, and dogs, horses and pigs were found to have low risk for ACE2 binding.
How this relates to infection and disease risk needs to be determined by future studies, but for those species that have known infectivity data, the correlation is high.
In documented cases of SARS-COV-2 infection in mink, cats, dogs, hamsters, lions and tigers, the virus may be using ACE2 receptors or they may use receptors other than ACE2 to gain access to host cells.
Lower propensity for binding could translate to lower propensity for infection, or lower ability for the infection to spread in an animal or between animals once established.
Because of the potential for animals to contract the novel coronavirus from humans, and vice versa, institutions including the National Zoo and the San Diego Zoo, which both contributed genomic material to the study, have strengthened programs to protect both animals and humans.
“Zoonotic diseases and how to prevent human to animal transmission is not a new challenge to zoos and animal care professionals,” said co-author Klaus-Peter Koepfli, senior research scientist at Smithsonian-Mason School of Conservation and former conservation biologist with the Smithsonian Conservation Biology Institute’s Center for Species Survival and Center for Conservation Genomics.
“This new information allows us to focus our efforts and plan accordingly to keep animals and humans safe.”
The authors urge caution against overinterpreting the predicted animal risks based on the computational results, noting the actual risks can only be confirmed with additional experimental data. The list of animals can be found here.
Research has shown that the immediate ancestor of SARS-CoV-2 likely originated in a species of bat. Bats were found to be at very low risk of contracting the novel coronavirus via their ACE2 receptor, which is consistent with actual experimental data.
Whether bats directly transmitted the novel coronavirus directly to humans, or whether it went through an intermediate host, is not yet known, but the study supports the idea that one or more intermediate hosts was involved.
The data allow researchers to zero in on which species might have served as an intermediate host in the wild, assisting efforts to control a future outbreak of SARS-CoV-2 infection in human and animal populations.
Virus jumping the species barrier, zoonotic spillover, transmission to humans
Virus jumping the species barrier
he SARS-CoV-2 has first been reported from the pneumonia patients of the Wuhan city in Hubei province of China. These patients were involved in trading at a wet animal market in the Huanan area. It is believed that SARS-CoV-2 is introduced from the animal kingdom to human populations during November or December 2019, as revealed from the phylogeny of the genomic sequences from the initially reported cases [94].
The spillover of SARS-CoV-2 from animals to humans took place at the beginning of December 2019 [56], and the clinical cases appeared around ending December [46,95].
Genetic analysis showed that this novel virus is closely related to bat CoVs and is similar but distinct from the SARS virus [43].
Several evidences based on genome sequences, the homology of the ACE2 receptor, and the presence of single intact ORF on gene 8 indicate bats as a natural reservoir of these viruses.
However, an unknown animal is yet to be unravelled as an intermediate host [43,46,47,49,56]. Initial investigations on animal source origin of SARS-CoV-2 have inconclusively revealed snakes [27], pangolins, and turtles [96].
The rapid spread of COVID-19 followed the initial animal to human spillover through human-to-human transmission. Genetic epidemiology had revealed that the spread from the beginning of December when the first cases were retrospectively traced in Wuhan was mainly by a human-to-human transmission and not due to continued spillover [56].
These species cross jumping, spillover, and rapid transmission events are linked to viral characteristics, host diversity, and environmental feasibility.
Coronaviruses being RNA viruses have high mutation rates that, besides creating new strains, enable them to adapt to a wide range of hosts. Hence, based on genome sequences, all known human CoVs have emerged from animal sources [97].
This seventh member of the human CoV has also been isolated initially from the pneumonia patients who were having direct or indirect links to the Huanan seafood market in Wuhan China, wherein other animals were also being sold [98].
These include a 49-year-old lady retailer in this wet animal market, a 61-year-old frequent visitor to this market, and a 32-year-old man [74,98]. Further, isolation of the SARS-CoV-2 from the environmental samples around this market, including people, animals, soil, discharges, or structures, strengthens the claims of involvement of hosts either as a reservoir or intermediate [26,52,99].
Recently, a Pomeranian dog as a probable intermediate host was identified; however, such reports are yet to be validated, and research is underway to explore the emergence of this infectious disease at the animal-human interface [29,99].
Multiple substitutions were observed in ACE2 receptors of a dog [96]. In this context, the Pomeranian dog of the infected owner found positive for COVID-19 suggest the permissiveness of the species for SARS-CoV-2 as a result of species jumping [99,100].
Among the fifteen dogs tested from different households with confirmed human COVID-19 cases in Hong Kong SAR, two dogs were found to be infected with SARS-CoV-2. The diagnosis was made using quantitative RT–PCR, serology, and viral genome sequencing.
Virus isolation was also done from the samples obtained from one dog. The genetic sequences of viruses obtained from the two dogs were identical to the ones that were detected from their human cases indicating human-to-dog transmission [101].
Moreover, a study reported that the SARS-CoV-2 might infect the cats and further transmitted by the infected cat to other cats [102]. One cat was tested positive for SARS-CoV-2 in France that showed mild respiratory and digestive signs.
The cat was tested positive by RT-qPCR on the rectal swab, and serological analysis identified the presence of antibodies against SARS-CoV-2. Genome analysis further confirmed that the SARS-CoV-2 isolated from the cat belongs to the phylogenetic clade A2a seen in French human indicating human-to-animal transmission [103].
This is not the first time that a domestic cat has been found susceptible to zoonotic coronavirus. During the 2003 SARS-CoV outbreak, domestic cats were tested positive for SARS-CoV that were living near SARS infected humans [21,104].
Even though experimental evidence indicates the possibility of SARS-CoV-2 transmission from infected to a susceptible cat close, SARS-CoV-2 transmission between cats or cat-to-humans are not reported under natural conditions [21].
Furthermore, along with dogs and cats, the zoo animals like tigers and lions were also reported to get the SARS-CoV-2 infection and exhibit clinical signs such as vomiting, diarrhoea, dry cough, breathing difficulty and wheezing [105,106].
Spillover of SARS-CoV-2 was also reported in mink farms of Netherlands, further increasing the concern of transmission to humans. Outbreaks of SARS-CoV-2 were reported in two mink farms holding 12,000 and 7500 animals. The virus is suspected to be introduced by a farmworker having COVID-19 [107,108].
Host-pathogen interactions and pathogenesis determine the severity and expression of disease [30,[109], [110], [111]]. Adaptation over time reduces the severity of infection as happened with HCoVs; however, the emergence of novel viruses or strains due to genetic alterations or recombinations can enhance hardness producing novel diseases like COVID-19 [111,112].
Evolutionarily, the balance of viral-human interaction and immune response against virus enables adaptation, thereby persistence in a host without severe or symptomatic disease when the aggravated pathogenesis results in mortality. Hence, loss of sustainable hosts and transmission to novel hosts becomes inevitable for future sustainability [111,112].
A pathogen cannot kill all its hosts, and for future sustainability, it adapts to some suitable host or spills over to a new host.
Zoonotic spillover
SARS-CoV-2 has been implicated to be originated from animals, and associated with animal linkages, spillover events, cross-species barrier jumping and zoonosis [9,20,27,[29], [30], [31],113].
Since the beginning of 2002 till the end of 2019, three coronaviruses viz. SARS-CoV, MERS-CoV, and SARS-CoV-2 have caused havoc in the human population globally and will continue to do so.
Earlier identified betacoronaviruses (SARS-CoV and MERS-CoV) were reported in Guangdong province of China in November 2002 and Saudi Arabia in 2012, respectively [114].
SARS-CoV-2 is the third zoonotic betacoronaviruses recognized in this century.
However, the CFR of the SARS-CoV-2 is lower to date when compared with SARS and MERS. It should not be overlooked as many asymptomatic cases may remain undiagnosed due to the unavailability of diagnostic kits in China.
With nearly 0.62 million deaths till the preparation of the manuscript, SARS-CoV-2 is proven to be deadliest as far as the number of deaths is concerned in comparison with SARS-CoV and MERS-CoV with 774 and 858 associated deaths, respectively [115,116].
Earlier, COVID-19 was linked with the exposure to the Huanan seafood market. However, individuals with no history of exposure above were also diagnosed with the illness, further supporting the human to human spread through droplets produced by cough and sneeze [66].
The spread of COVID-19 that occurred with a high pace and lack of transparency in reporting the disease by the Chinese Health Ministry and failure in the timely implementation of preventive measures has been considered as the primary contributor as stated earlier in SARS [19,117].
Both SARS-CoV and SARS-CoV-2 showed prominent similarities in their pathogenesis and epidemics. In both cases, bats were considered as the natural host, and the cold temperature and low humidity in cold, dry winter provided conducive environmental conditions that promoted the survival of the virus in the environment [62].
Further, Moriyama et al. [118] assessed the significance of the environmental factor on host immune system targeting innate and adaptive both responses in the respiratory tract.
Zoonotic spillover is the transmission of pathogens to humans from vertebrate animals [119]. At present, these spillovers are of significant concern as in the past, many spillovers in the form of Nipah, Hendra, Ebola, SARS, MERS, and ongoing COVID-19 involving many animal species like pigs, horses, monkeys, camels, civets, among others, were documented.
Bovine CoVs have been reported to infect children and thus possess zoonotic potential [9,13,120,121]. Spillover is governed by the interaction of viral-specific proteins like S protein and host ACE2 receptor [30,31,111,112].
These S proteins have RBD in CoVs, which contain receptor binding motifs (RBM) that help in specific binding to host ACE2 receptors [47,122]. Mutations in amino acid sequences of RBDs results in a change in specificity of a receptor, interaction and binding, hence alteration in transmissibility, pathogenicity and cross-species jumping with a predisposition to novel and more severe diseases [110,123].
In the case of SARS-CoV-2, RBD of S protein has 10–15 times affinity ACE2R [88,110]. It has furin recognition sequence “RRAR” at the S1–S2 cleaving site that represents a functional site for the cellular serine protease TMPRSS2 thus increasing the efficiency of transmission and contagiousness [88,90,110].
In addition to enhanced binding affinity, electrostatic complementarity and hydrophobic interactions are critical to enhancing receptor binding and escaping antibody recognition by the RBD of SARS-CoV-2, thereby further increasing transmission capability and contagiousness [110].
A detailed investigation regarding the emergence of new coronavirus, host range, and transmissibility is crucial to understand such pandemics shortly. The literature revealed that before the appearance of SARS-CoV and MERS-CoV, human coronavirus (HCoV) strains like HCoV-NL63, HCoV-229E, HCoV-OC43, and HCoV-HKU1 were the CoVs strains producing mild infections in humans.
However, their natural ancestral hosts were of animal origin, like bats for HCoV-NL63, and HCoV-229E and rodents were natural hosts for HCoV-OC43 and HKU1. These four HCoVs were initially of low pathogenicity. To enhance the pathogenicity, they used intermediate hosts such as cattle for HCoV-OC43 (natural host was rodent), and alpacas for HCoV-229E (bats were natural host) and this way acquired the ability to infect human beings with serious health hazards [124].
Added to the involvement of bats and pangolins, the recent reports revealing SARS-CoV-2 infection in cats, dogs, tigers, lions and minks have raised concerns over this virus affecting multiple animal species, and also points out towards the incidences of reverse zoonosis [31,102,106,108,125,126].
The ferrets, cats, and primates are suggested to be good candidates for susceptibility to SARS-CoV-2 [123,127].
COVID-19 research and surveillance in companion and pet animals, livestock animals, zoo animal species, wildlife animal species as well as their handlers, veterinarians, and owners need to be enhanced during the pandemic, which would help to follow better integrated one health strategies [128] and appropriate preventive and mitigation to counter SARS-CoV-2 effectively [29,126,[129], [130], [131], [132], [133]].
Significance of COVID-19 monitoring and implementation of suitable public health measures among workers involved in meat and poultry processing facilities/industries has been emphasized, which would protect them as well as aid in preserving the critical meat and poultry production infrastructure and the meat products [134].
Transmission to humans
The involvement of intermediate hosts in maintaining and transmitting the virus to susceptible host predisposes humans to novel CoVs leading to the emergence of new diseases in humans.
The currently ongoing SARS-CoV-2/COVID-19 pandemic has put on hold the entire world [111,135]. The CoVs have frequently been associated with animal and human diseases and have a zoonotic interface [97,111].
Usually, one or more types of animal hosts are involved in the transmission cycle of CoVs to humans [30,97]. That can be natural host, reservoir host, intermediate host or definitive host [31].
Bats have been the natural hosts for human CoVs of Alphacoronavirus (HCoV-NL63, HCoV-229E) and Betacoronavirus (SARS-CoV, MERS-CoV, SARS-CoV-2) genera whereas for Betacoronavirus members HCoV-OC43 and HCoV-HKU1, rodents are the natural hosts.
Genome sequence analysis has revealed bats as a natural host for SARS-CoV-2 [30,97]. In natural or reservoir hosts, CoVs adapts well, however, being unstable RNA viruses, they keep multiplying continuously without producing disease thereby enabling persistence or survivability and accumulation of mutations over the time resulting in the emergence of newer and novel strains of viruses [97,111,136].
These unique strains or viruses occasionally spill over to other species including animals or humans, adapting to their body systems and hence broaden the biological host range for evolutionary sustainability; however, results in epidemiological widening of disease sphere as well [25,31].
This transmission and adaptation scenario initiates a host-pathogen response resulting in the novel usually severe diseases that can at times be fatal in initial stages or over extended periods until virus pathogen adapts to host or the host develops sufficient immune defence [137,138].
It has been reported that almost all HCoVs have originated from animals like bats (SARS-CoV, MERS-CoV, HCoV-NL63, and HCoV-229E) and rodents (HCoV-OC43 and HKU1) [139,140]. Additionally, CoVs have been reported to infect several species of domestic and wild animals either clinically or subclinically [27,28,30,141].
Cattle, horses, camels, swine, dogs, cats, birds, rabbits, rodents, ferrets, mink, bats, snakes, frogs, marmots, hedgehogs, Malayan pangolin along with other wild animals may serve as a reservoir host of coronavirus [9,27,44,124,[142], [143], [144], [145], [146], [147]].
In the context of SARS-CoV-2, snakes, pangolins and bats have been suspected as intermediate hosts since the first cases of COVID-19 had links to Huanan Sea Food Market where different animals, birds, and wild animals were being sold along with seafood items [27,28,43,109,147,148].
Coronaviruses have been reported to cause salivary, enteric and respiratory infections in laboratory animals (mice, rat, guinea pig, and rabbit) and urinary tract infection, respiratory illness and reproductive disorder in poultry [9,142].
In bovine, canine, feline and swine CoVs infections have resulted in diarrhoea, enteritis, respiratory illness, gastro-intestinal affections and nervous symptoms [120,121,[149], [150], [151]].
Coronavirus, namely- SW1, has been reported in captive beluga whale using a panviral microarray method [152]. Among all the assumptions on animal hosts as the intermediate host, genomic and evolutionary information from pangolins reveals the highest closeness to the SARS-CoV-2 than any other host CoVs isolates [153].
The spike protein, the main target of many studies searching for a cure of COVID-19, has been found highly similar to SARS-CoV-2 and, thus, could serve as a surrogate system for further evaluations [153].
Bats are the natural reservoir host of many CoVs. As reported earlier, 7 out of 11 alphacoronaviruses and 4 out of 9 betacoronaviruses as per the International Committee on Taxonomy of Viruses (ICTV) classification were solely originated from bats [97,154].
According to the literature, bats have been regarded as a potential wildlife reservoir whereas civets and dromedary camels as intermediate hosts of SARS-CoV and MERS-CoV, respectively [155,156].
The bat coronavirus, BatCoV RaTG13, has shown higher relatedness to SARS-CoV-2 at the whole genome level and spike gene in particular [153]. Coexistence and frequent recombination between highly diversified and prevalent bat SARS-related coronaviruses (SARSr-CoV) and coronaviruses may suggest the probable emergence of novel viruses shortly [157,158].
Benvenuto et al. [159] analyzed the whole genome sequences of different CoVs using Fast Unconstrained Bayesian AppRoximation (FUBAR) to understand the evolutionary and molecular epidemiology of SARS-CoV-2.
The authors concluded that SARS-CoV-2 clustered with sequences of bat SARS-like CoVs with a few mutations in nucleocapsid and spike glycoprotein, suggesting its probable transmission from the bats [159].
Bats, especially horseshoe bats (Rhinolophus spp.), are considered to be the known reservoirs of SARS-related CoVs. Since the bat origin, CoVs have always caused outbreaks in humans, studying the diversity and distribution of coronavirus populations in the bats will help to mitigate future outbreaks in humans and animals [160].
Interestingly, bats play a crucial role in all the spillovers mentioned above, indicating their importance in the emergence of new viruses. The reason behind the emergence and broad host range of CoVs in the past and present might be due to unstable RNA-dependent RNA polymerase (RdRp), lack of proof-reading ability, high frequency of mutations in the receptor-binding domain of spike gene and genetic recombination [140,161,162].
Bat CoVs have high diversity and great potential of spillover in different animal species, as reported earlier in civet cat and dromedary camel, leading to well-known pandemics SARS and MERS, respectively along with the recent spillover in pigs resulted in swine acute diarrhoea syndrome (SADS).
However, spillover resulted in the emergence of SADS-CoV, which showed a 95% genomic identity with bat coronavirus, which led to severe mortality with 24,693 deaths in neonatal piglets [160].
Fortunately, it did not excel in the form of the third pandemic, and no human cases were reported till date. The spillover responsible for ongoing COVID-19 is still under investigation and a matter of great concern for the researchers all around the globe.
Based on Resampling Similarity Codon Usage (RSCU), snakes (Bungarus multicinctus and Naja atra) were suggested as wildlife reservoirs of SARS-CoV-2 and reported to be associated with the cross-species transmission [27] and later it was disapproved by other researchers [163,164].
Unfortunately, to date, the intermediate host of the SARS-CoV-2 is abstruse what results in its escalation in the human population around the globe.
In this context, analyzing the interaction between the Asn501 site in RBD of spike glycoprotein of SARS-CoV-2 and the residue at 41 sites of ACE2 receptor of different hosts (pangolins, turtle, mouse, dog, cat, hamster and bat) revealed that tyrosine has higher receptor binding affinity than histidine suggesting pangolins and turtle be closer than bats to humans and maybe the probable intermediate hosts of SARS-CoV-2 [96].
However, this hypothesis was also contradicted by Li et al. [28] based on an insertion of the unique peptide (PRRA) in the SARS-CoV-2 virus, which was lacking in CoVs from pangolins.
Moreover, SARS-CoV-2 showed higher similarity to the BetaCoV/bat/Yunnan/RaTG13/2013 compared to the ones that were isolated from the pangolins, thereby denied the direct link of the virus from pangolins. However, further studies are required to confirm the role of pangolins in SARS-CoV-2 spread to humans.
The receptor-binding domain of the spike protein of SARS-CoV interacts with the host receptor ACE2 facilitating its potential of cross-species, as well as human-to-human transmission [47]. Similarly, the spike protein of SARS-CoV-2 was reported to recognize ACE2 receptors expressed in fish, amphibians, reptiles, birds, and mammals and has a more robust binding capacity (affinity) in comparison to SARS-CoV [89].
This suggests their involvement as probable natural and intermediate hosts [161], which may further help in the selection of animal models for epidemic investigation and preventing its spread [47].
Bat origin CoVs have been found to cross the species barrier that favoured their transmission via recombination/mutations in the RBD. The evidence of a virus outbreak that occurred in Chinese pig farms suggests its possible cross-species [160,165].
Also, murine cells were found permissive for SARS-CoV after substitution of His353 with Lys353 in the ACE2 receptor of a mouse, which suggests the role of residue changes in the cross-species and human-to-human transmission [166].
Mutation in residues at position 479 and 487 of receptor binding motif (RBM) of SARS-CoV was reported to play a role in civet-to-human and human-to-human transmission, respectively [167,168].
The CoVs are more prone to recombination and mutations leading to variable host range, and resemblance of receptors in various hosts results in cross-species jumping [31,112,169]. Genetic divergence due to these genetic alterations results in the evolution of newer viral strains having altered virulence, tissue tropism, and host range [170,171].
Moreover, the presence of threonine at position 487 was reported to enhance the binding affinity of RBM for the ACE2 receptor of civet and humans [172]. However, many SARS-related coronaviruses (SARSr-CoV) have been reported in bats and used ACE2 receptors for entry into a host cell, which showed its potential to infect humans directly without any intermediate host [173].
In addition to this, no direct transmission of SARSr-CoV is reported from bats to humans to date. However, seropositivity on a serological investigation of individuals without prior exposure to SARS-CoV residing near bat caves in China revealed likely infection of humans by bat SARSr-CoV and related viruses [174]. Besides, the interspecies transmission potential of SARSr-CoVs is due to the ORF8 gene [175].
As per reports, the SARS-CoV emerged via recombination of bat SARSr-CoVs, was transmitted to farmed civets along with other mammals, and these infected civets spread the virus to market civets.
The virus was reported to undergo mutations in infected market civets before its spillover to humans. Similarly, the MERS-CoV circulated for 30 years in camels before the pandemic [97,176] supporting the hypothesis that after species jumping the exogenous viruses opted for adaptation to the environment and host before spillover to humans [177].
Moreover, the possible spillover of other circulating bat SARSr-CoVs to humans from mammalian hosts soon is highly anticipated.
The cross-species jumping and adaptation are determined by the presence of specific receptors on host tissues (like ACE 2 receptor for HCoV-NL63, SARS-CoV and SARS-CoV-2, dipeptidyl peptidase-4 for MERS-CoV, human aminopeptidase N for HCoV-229E, 9-O-acetylsialic acids for HCoV-OC43, HCoV-HKU1) which help in binding and entry of the virus into host cells [30,31].
These receptors are present in various body systems in animals and humans, including respiratory and gastrointestinal systems [30]. Reservoir host animals including bats and rodents possess these receptors which are similar to those present in camels, masked palm civets (Paguma larvata), or bovines, that act as an intermediate host for different CoVs [30,110,111].
Presence of some of these receptors in humans like ACE2 or DPP4 makes them vulnerable to CoV infection like SARS-CoV and MERS-CoV causing SARS and MERS infections, respectively [31,168].
The MERS-CoV spike was found to possess the capacity for adapting to species variation in the host receptor DPP4 [37]. The mechanism expressed by MERS-CoV in adapting to infect cells of new species might be present in the other coronaviruses.
ACE2 has also been found as a binding receptor for SARS-CoV-2 [47]. The species-specific variations in the host receptors limit the interaction with CoV spike protein, and this is responsible for the development of the species barrier that prevents spillover infection.
Snakes, civets, and pangolins are considered as the potential intermediate hosts of COVID-19. However, further confirmation is required by tracking the origin of the virus. This is critical for preventing additional exposure to this fatal virus [178].
The probability of the SARS-CoV-2 spread during incubation and convalescent period has been suggested [76]. As per reports, presence of CoVs has been observed in respiratory droplets, body fluids and inanimate objects with the ability to remain infectious for nine days on contaminated surfaces resulting in its risk of self-inoculation via mucous membranes of the eyes, mouth or nose [[179], [180], [181]].
Nosocomial, as well as human-to-human transmission, have been reported to occur via virus-laden aerosols, contaminated hands or surfaces, and close community contact with an infected person [66,182,183].
The ocular route has been reported in the human-to-human transmission of SARS-CoV-2, as observed in SARS-CoV, suggesting the involvement of different ways other than the respiratory tract [184,185].
Later on, the probability of the faecal-oral route for potential transmission of the virus was also suggested [186]. The metatranscriptome sequencing of SARS-CoV-2 in the bronchoalveolar lavage fluid (BALF) of infected individuals resulted in polymorphism in few intra-hosts variants, suggesting the in vivo evolution of the virus thereby affecting its virulence, transmissibility, and infectivity [187].
An overview of coronaviruses jumping the cross-species barriers, zoonotic CoVs transmitted from bats to animals before spillover to humans, and possible prospects for further transmission to mammalian hosts is depicted in Fig. 1 .

Cross-species transmission of known zoonotic coronaviruses from bats to animals before spillover to humans and probable prospects of further transmission to mammalian hosts.
reference link : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7396141/
More information: “Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates,” Proceedings of the National Academy of Sciences (2020). DOI: 10.1073/pnas.2010146117 , www.pnas.org/content/early/2020/08/20/2010146117



{kind=link}