










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



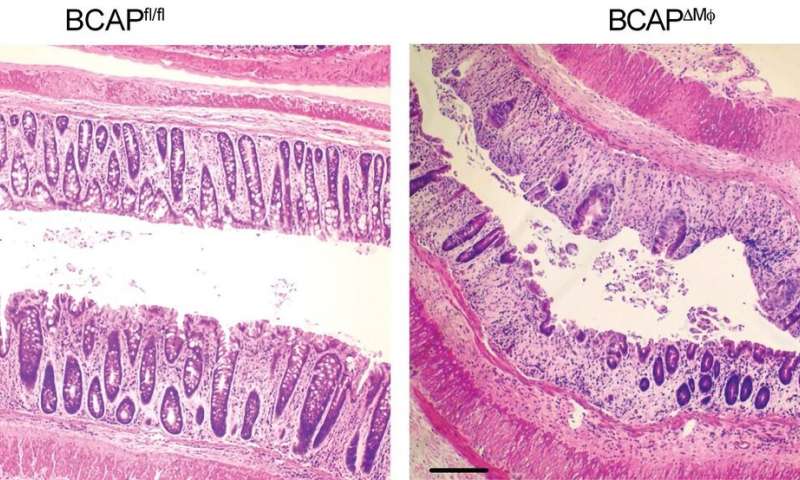
Managing the activity of the BCAP protein could help the body repair intestinal tissue from damage caused by inflammation, according to a new study led by experts at Cincinnati Children’s.
The findings, published online Nov. 16, 2020, in PNAS, suggest that targeting BCAP with new medications could benefit people living with conditions such as inflammatory bowel disease (IBD) and rheumatoid arthritis.
The multi-center research team was led by first author Ricardo Irizarry-Caro, BSc, and senior author Chandrashekhar Pasare, DVM, Ph.D., at Cincinnati Children’s Division of Immunobiology,
“This study provides fundamental understanding of how macrophages regulate their responses to pathogens to provide an effective yet controlled inflammatory response and avoid damage of the host tissue,” Pasare says.
“More importantly, this study has implications for small-molecule drug discovery targeting BCAP, which could be used to either promote reparative macrophage transition or induce a robust inflammatory response by just altering one single protein.”
Dampening inflammation would be helpful for managing inflammatory diseases such as IBD and rheumatoid arthritis.
Meanwhile, in cancer treatment, promoting inflammation could help drive immune responses against solid tumors.
Macrophages and Inflammation
When macrophages sense bacterial pathogens such as E.coli, they activate an inflammatory response to get rid of the pathogen. However, this response needs to be kept in check to avoid damaging host tissue.
The new study shows that macrophages employ a molecule called BCAP to transition into a reparative state following clearance of infection.
Turning down macrophage-triggered immune response once their task is complete is important because uncontrolled inflammation can lead to diseases such as inflammatory bowel disease (IBD).
Macrophages utilize Toll-Like Receptors (TLRs) to sense various pathogens, and activation of these receptors leads to early inflammation.
The authors found that downstream of TLR activation, BCAP simultaneously activates a parallel pathway called the PI3K-AKT pathway, which acts to limit these inflammatory signals.
Specifically, researchers found that BCAP downstream of TLR activation inhibited two key proteins, GSK3b and FOXO1, which places a cap on the inflammation triggered by pathogen sensing. Yet this cap was not enough by itself to make macrophages stand down.
The team also found that BCAP was necessary for promoting a metabolic process called ‘histone lactylation,’ which leads to macrophage transition and supports intestinal repair after injury.
The co-authors say finding or developing drugs capable of controlling BCAP activity would be an important future goal for drug research.
ability (1, 2). Upon pathogen encounter, DCs undergo a process called “maturation,” characterized by the up-regulation of surface co-stimulatory receptors, major histocompatibility complexes (MHCs), and the production of pro-inflammatory cytokines, all of which are important for the priming of naive T cells (1, 3, 4).
A normal maturation of DCs is indispensable for the establishment of powerful innate and adoptive immune responses during infection, and abnormal maturation of DCs is often linked with immune disorders and diseases. While excessive maturation of DCs leads to transplant rejection and the onset of autoimmune disease (5, 6), loss of normal DC maturation results in uncontrollable infection and inflammation.
It has become clear that immature DCs mainly elicit immunological tolerance and are critical for the establishment of peripheral tolerance (5, 7, 8), which further underlines the critical role of mature DCs in shaping appropriate adaptive immune responses.
The maturation of DCs is mainly caused by the activation of pattern recognition receptors (9–11), the best-characterized of which are the Toll-like receptors (TLRs). Activation of TLRs induces a series of intracellular signaling cascades including the NF-κB, MAPK, and PI3K/AKT signaling in a MyD88- dependent and/or TRIF-dependent manner (9, 12).
TLR- mediated activation of NF-κB and MAPK signaling is responsible for initiating the transcription of pro-inflammatory cytokines and co-stimulatory receptors. The activation of PI3K/AKT signaling in DCs acts mainly as a negative regulator of TLR signaling (13, 14).
The activation and crosstalk of these signaling networks are delicately monitored and still require more investigation.
BCAP acts as a signaling adaptor and contains numerous potential protein-protein interaction domains, but does not contain any enzymatic domains (15). Through protein-protein interactions, BCAP can orchestrate multiple intracellular signals downstream of receptor and non-receptor tyrosine kinases. BCAP was initially described as an adaptor for PI3K/AKT signaling transduction downstream of BCR and CD19 in B cells (16, 17).
BCAP-activated PI3K/AKT signaling is indispensable for B cell development and activation (18) but inhibits the activation of macrophages and NK cells (19, 20). These paradoxical results suggest that BCAP plays different roles in cell-type specific fashion. In addition to the PI3K/AKT signaling, BCAP also modulates the activation of NF-κB signaling.
BCAP- deficient mice have a reduced number of mature B cells due to impacted NF-κB signaling in B cells, suggested a non- redundant role of BCAP in maintaining the NF-κB signaling activation during B cell development (21). However, recent studies demonstrated that BCAP limits the activation of NF-κB signaling by interacting with TIR domain-containing proteins in macrophages (22, 23).
Although a recent report demonstrates that BCAP promotes IFN-α production through TLR7/9 signal transduction in pDCs (24), and BCAP has been found to be expressed in cDCs (22), its role in cDCs is yet to be elucidated. These results further highlight the diversity functions of BCAP, as it has the ability to interact with different effector proteins depending on the context.
Here, we describe a role of BCAP as a modulator of cDC maturation and conclude that BCAP acts as a negative regulator of TLR signaling through orchestrating the activation of NF- κB and PI3K/AKT signaling. This dual-regulation properties of BCAP relies on its bilateral interaction with MyD88 and p85α (a subunit of PI3K).
Consistently, BCAP impairs TLR-induced cDC maturation, limiting the adoptive immune responses against Listeria monocytogenes in mouse model. Collectively, these findings have uncovered a previous unknown strategy for the regulation of cDC maturation in mice.
REFERENCES
- Steinman RM. Decisions about dendritic cells: past, present, and future. Annu Rev Immunol. (2012) 30:1–22. doi: 10.1146/annurev-immunol-100311-102839
- Iwasaki A, Medzhitov R. Control of adaptive immunity by the innate immune system. Nat Immunol. (2015) 16:343–53. doi: 10.1038/ni.3123
- Roche PA, Furuta K. The ins and outs of MHC class II-mediated antigen processing and presentation. Nat Rev Immunol. (2015) 15:203–16. doi: 10.1038/nri3818
- Zhou J, Lai W, Yang W, Pan J, Shen H, Cai Y, et al. BLT1 in dendritic cells promotes Th1/Th17 differentiation and its deficiency ameliorates TNBS-induced colitis. Cell Mol Immunol. (2018) 15:1047–56. doi: 10.1038/s41423-018-0030-2
- Guerder S, Joncker N, Mahiddine K, Serre L. Dendritic cells in tolerance and autoimmune diabetes. Curr Opin Immunol. (2013) 25:670–5. doi: 10.1016/j.coi.2013.10.004
- Liu Q, Rojas-Canales DM, Divito SJ, Shufesky WJ, Stolz DB, Erdos G, et al. Donor dendritic cell-derived exosomes promote allograft-targeting immune response. J Clin Invest. (2016) 126:2805–20. doi: 10.1172/JCI84577
- Ganguly D, Haak S, Sisirak V, Reizis B. The role of dendritic cells in autoimmunity. Nat Rev Immunol. (2013) 13:566–77. doi: 10.1038/nri3477
- Li H, Shi B. Tolerogenic dendritic cells and their applications in transplantation. Cell Mol Immunol. (2015) 12:24–30. doi: 10.1038/cmi.2014.52
- Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. (2010) 11:373–84. doi: 10.1038/ni.1863
- O’Neill LA, Golenbock D, Bowie AG. The history of Toll-like receptors— redefining innate immunity. Nat Rev Immunol. (2013) 13:453–60. doi: 10.1038/nri3446
- Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell.(2010) 140:805–20. doi: 10.1016/j.cell.2010.01.022
- Zhou Y, Wu J, Liu C, Guo X, Zhu X, Yao Y, et al. p38α has an important role in antigen cross-presentation by dendritic cells. Cell Mol Immunol. (2018) 15:246–59. doi: 10.1038/cmi.2016.49
- Fukao T, Tanabe M, Terauchi Y, Ota T, Matsuda S, Asano T, et al. PI3K- mediated negative feedback regulation of IL-12 production in DCs. Nat Immunol. (2002) 3:875–81. doi: 10.1038/ni825
- Troutman TD, Bazan JF, Pasare C. Toll-like receptors, signaling adapters and regulation of the pro-inflammatory response by PI3K. Cell Cycle. (2012) 11:3559–67. doi: 10.4161/cc.21572
- Singh MD, Ni M, Sullivan JM, Hamerman JA, Campbell DJ. B cell adaptor for PI3-kinase (BCAP) modulates CD8(+) effector and memory T cell differentiation. J Exp Med. (2018) 215:2429–43. doi: 10.1084/jem.20171820
- Okada T, Maeda A, Iwamatsu A, Gotoh K, Kurosaki T. BCAP: the tyrosine kinase substrate that connects B cell receptor to phosphoinositide 3-kinase activation. Immunity. (2000) 13:817–27. doi: 10.1016/S1074-7613(00)00079-0
- Inabe K, Kurosaki T. Tyrosine phosphorylation of B-cell adaptor for phosphoinositide 3-kinase is required for Akt activation in response to CD19 engagement. Blood. (2002) 99:584–9. doi: 10.1182/blood.V99. 2.584
- Yamazaki T, Takeda K, Gotoh K, Takeshima H, Akira S, Kurosaki T. Essential immunoregulatory role for BCAP in B cell development and function. J Exp Med. (2002) 195:535–45. doi: 10.1084/jem.20011751
- MacFarlane AW, Yamazaki T, Fang M, Sigal LJ, Kurosaki T, Campbell KS. Enhanced NK-cell development and function in BCAP-deficient mice. Blood. (2008) 112:131–40. doi: 10.1182/blood-2007-08-107847
- Ni M, MacFarlane AW, Toft M, Lowell CA, Campbell KS, Hamerman JA. B- cell adaptor for PI3K (BCAP) negatively regulates Toll-like receptor signaling through activation of PI3K. Proc Natl Acad Sci USA. (2012) 109:267–72. doi: 10.1073/pnas.1111957108
- Yamazaki T, Kurosaki T. Contribution of BCAP to maintenance of mature B cells through c-Rel. Nat Immunol. (2003) 4:780–6. doi: 10.1038/ni949
- Troutman TD, Hu W, Fulenchek S, Yamazaki T, Kurosaki T, Bazan JF, et al. Role for B-cell adapter for PI3K (BCAP) as a signaling adapter linking Toll- like receptors (TLRs) to serine/threonine kinases PI3K/Akt. Proc Natl Acad Sci USA. (2012) 109:273–8. doi: 10.1073/pnas.1118579109
More information: Ricardo A. Irizarry-Caro el al., “TLR signaling adapter BCAP regulates inflammatory to reparatory macrophage transition by promoting histone lactylation,” PNAS (2020). www.pnas.org/cgi/doi/10.1073/pnas.2009778117



{kind=link}