










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


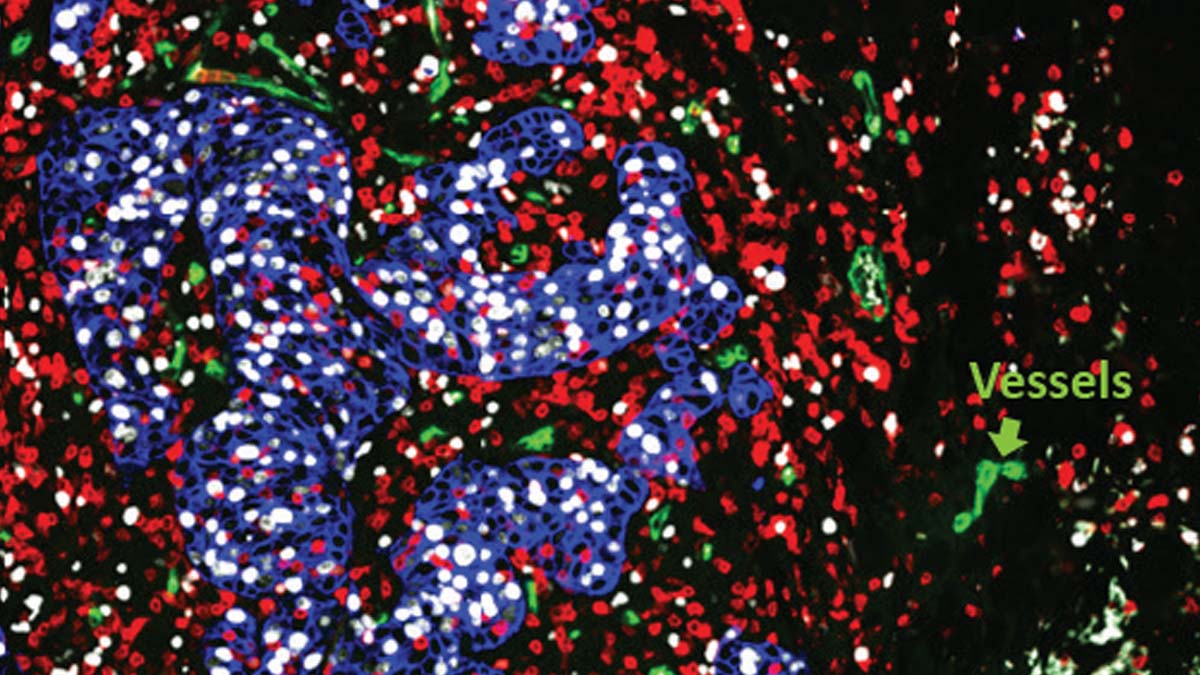
New research that is examining the “tumor microenvironment” reveals not only how macrophages can become extraordinary turncoats but also how they can actively support tumor growth and metastatic progression in certain forms of breast cancer.
The tumor microenvironment – or TME – refers to the region encompassing a tumor, an area entangled with blood vessels that feed the tumor, and aided by a support cast of immune cells, signaling molecules, fibroblasts, resident host cells, lymphatics, and an array of proteins.
Macrophages in the tumor microenvironment aren’t the faithful soldiers of the immune system gobbling up invaders. Instead, they’re shady traitors that aid and abet the enemy – cancer.
Tumors of all kinds actively engage with their microenvironment, a factor that strongly influences tumor progression and metastasis. A growing body of evidence suggests that triple negative breast cancer is among the forms of the disease that may benefit from research into the tumor microenvironment.
This form of breast cancer lacks all of the receptors for which treatments have been developed. Receptors are proteins that stipple the surface of cells, and certain drugs target these receptors to control the disease. Common proteins on breast cancer cells include HER2, estrogen and progesterone receptors.
Medical investigators are only now developing a clearer concept of the myriad ways in which breast cancer can proliferate and resist existing therapies. Studies involving the tumor microenvironment are opening a new window of understanding.
As it turns out, macrophages in the tumor microenvironment serve multiple roles in cancer progression and have become a potential prognostic factor for breast cancer.
Metabolic changes due to “cross-talk” between the tumor and its surrounding microenvironment is currently considered one of the emerging hallmarks of several types of malignancies, including breast cancer.
Dr. Valentí Gómez of University College London and a team of cancer biologists report in Science Signaling that a growth factor secreted by macrophages promotes metabolic changes in breast cancer cells. “Dysregulation of cellular metabolism constitutes one of the hallmarks of tumor progression,” Valenti and colleagues wrote.
“Because of their high proliferative rate, cancer cells have a high demand for both energy – adenosine triphosphate (ATP) – and biosynthetic precursors.”
Although most normal cells derive energy by using glycolysis as a first step before turning to a more sophisticated process called oxidative phosphorylation, cancer cells rely solely on glycolysis to generate energy. Glycolysis is a primordial form of energy production and was likely the type of energy production of the planet’s earliest forms of life.
It is a much less efficient process for extracting energy, but doesn’t require oxygen, which is limited in tumors, and probably is the reason that tumors rely so heavily on glycolysis, Gómez asserted.
In a series of experiments, Gómez and the London-based team, found that secretion of the cytokine TGF-β from anti-inflammatory tumor-associated macrophages reduced the abundance of succinate dehydrogenase, an enzyme that is critical for oxidative phosphorylation in breast cancer cells.
The loss of this metabolic enzyme promoted an increase in glycolysis, thereby enhancing tumor growth, blood vessel growth, and immunosuppression.
Macrophages in this dynamic microenvironment frequently support tumor growth and metastatic progression.
In experiments involving mice, Gómez and colleagues found that depleting anti-inflammatory tumor-associated macrophages or blocking TGF-β suppressed these effects in the animals. The findings revealed a metabolic mechanism underlying the tumor growth-promoting roles of tumor-associated macrophages, Gómez and colleagues found.
“We found that anti-inflammatory tumor-associated macrophages promoted a metabolic state in breast cancer cells that supported various pro-tumorigenic phenotypes,” Gómez wrote.
Even anti-inflammatory tumor-associated macrophages secreted the cytokine TGF-β that, upon engagement of its receptors in breast cancer cells, suppressed the abundance of specific transcription factors while consequently decreasing the metabolic enzyme succinate dehydrogenase in tumor cells.
When that happened – the decrease in succinate dehydrogenase levels – tumor cells accumulated succinate, which enhanced the stability of the transcription factor HIF1α and reprogrammed cell metabolism to a glycolytic state.
Research, like that in the Gómez lab – and beyond – is important because it sheds new light on why cancer treatments fail and the tumor itself becomes resistant to treatment.
“Pro-tumorigenic macrophages contribute to cancer progression by affecting the central glucose metabolism, angiogenesis, and immune evasion within the tumor, and their presence may partially explain the limited efficacy of antiglycolytic treatments,” Gómez explained.
New therapies are under development targeting specific constituents of the microenvironment.
“A novel combined approach should be evaluated to improve the outcome of triple-negative breast cancer therapies. For example, the use of nanoparticles to deplete/re-educate the … macrophage population within the tumor presents an intriguing therapeutic possibility, given their established association with poor prognosis.”
TAMs: General characterization
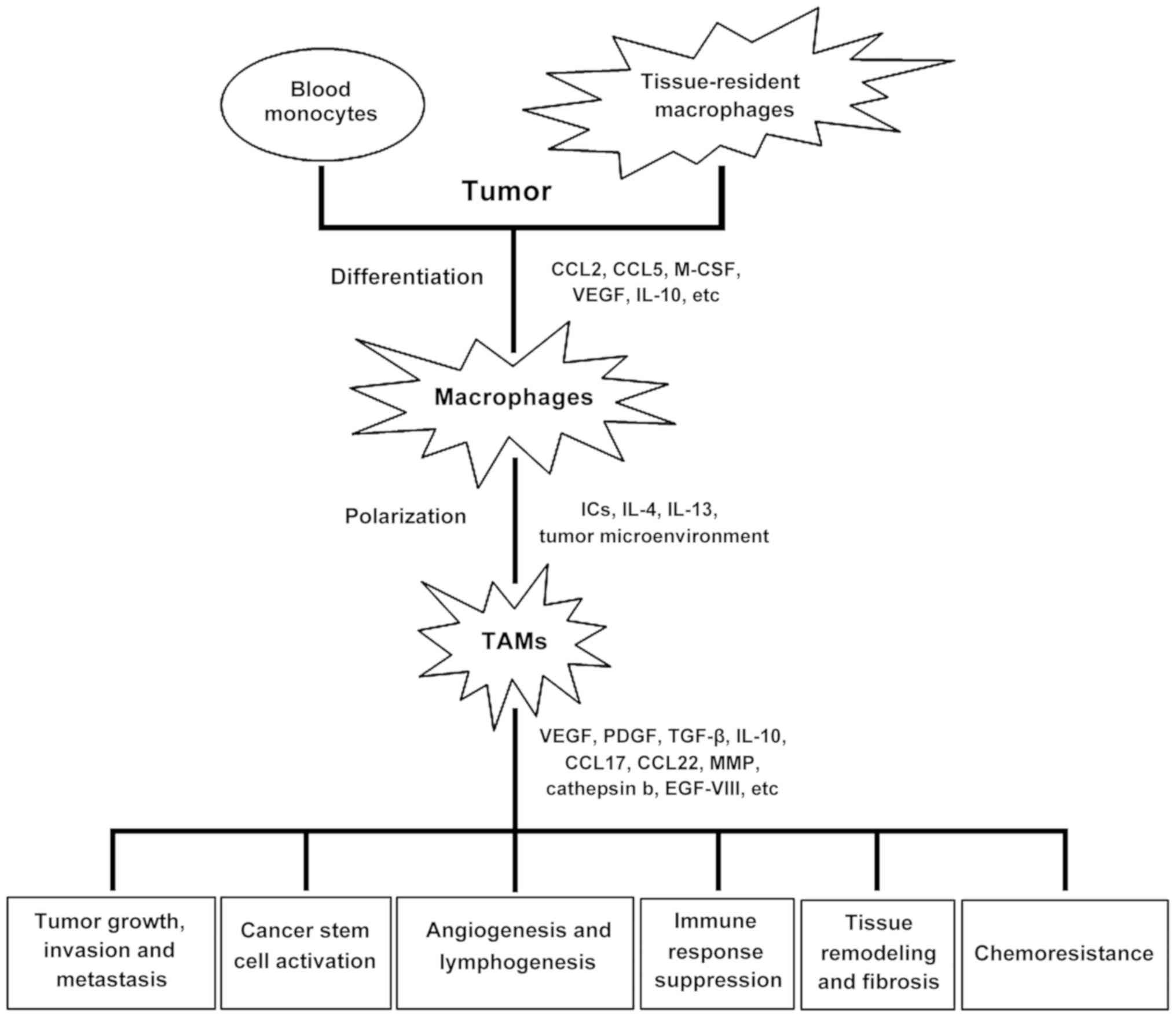
The process of tumorigenesis in the body begins to affect the tumor microenvironment, including macrophages. Blood monocytes penetrate the tumor, and differentiate into macrophages with an anti-inflammatory phenotype in response to signaling molecules produced by the tumor, such as interleukin (IL)-4, IL-10 and transforming growth factor (TGF)-β.
These signals suppress antitumor immunity, and stimulate the development of new blood vessels and thus tumor growth and metastasis (17).
The role of TAMs in tumor progression are illustrated in Fig. 1.
TAMs have attracted substantial attention for the past 30 years (from the time when the concept of a macrophage dichotomy was advanced) (25,26). TAMs are classed as type II-activated macrophages (M2). Stein et al (27) first characterized TAMs as alternatively activated macrophages in 1992.
Data on TAM markers and TAM-suppressing factors subsequently accumulated in further studies (28-31). The M2 population is highly heterogeneous (32,33). Macrophages with the M2 phenotype serve an important role in the process of tumorigenesis by suppressing the immune response, remodeling the extracellular matrix and stimulating angiogenesis (26).
M2 macrophages are characterized by the expression of specific receptors, such as arginase-1, mannose receptor (CD206), CD163, CD209, FIZZ1 and Ym1/2 (22,29).
Macrophages with the M1 phenotype (classically activated macrophages) express bactericide molecules and receptors (34). Macrophages acquire the M1 phenotype in response to endogenous inflammatory stimuli, such as the Th1-associated cytokine interferon-γ, or exogenous stimuli, such as lipopolysaccharides (23,35).
M1 macrophages produce pro-inflammatory cytokines and thereby stimulate the inflammatory response (36). A total of 5,598 publications on TAMs were available on PubMed as of July 10, 2020. The annual number of such publications increased from 51 in 2007 to 660 in 2019.
Macrophages are intricately involved in the immune response, and thus serve a protective role. They participate in the clearance of cellular debris and iron processing, degradation of dead cells and foreign material, response to infection, immunomodulation and modulation of inflammatory processes, angiogenesis, and facilitating wound healing (20,22,23).
Furthermore, macrophages serve an important in organ development, and in tissue turnover and regeneration (37,38). Adverse reactions are also often caused by macrophages and are associated with their M1/M2 polarization. M1 macrophages serve critical roles in innate host defense and in the killing of tumor cells.
Therefore, they are considered as antitumor macrophages. M2 macrophages tend to exert an immunosuppressive phenotype, favoring tissue repair, and tumor promotion. Thus, they are considered as pro-tumorigenic macrophages. The expression of inhibitory cytokines in tumor cells or macrophages provides a mechanism of resistance to anticancer therapy. Hence, a therapeutic strategy targeting macrophages or macrophage-derived cytokines may be a promising and effective method for targeting tumorigenesis (39).
TAMs are an important component of the tumor microenvironment, which affects tumor growth, tumor angiogenesis, immunity suppression, metastasis and chemoresistance. TAMs substantially affect the clinical efficacy of these drugs and drug resistance. For example, TAMs release chemoprotective factors, such as cathepsin b and milk-fat globule EGF-VIII, which promotes tumorigenicity of cancer stem cells and induces anticancer drug resistance.
Furthermore, drugs targeting TAMs have been shown to exhibit promising results for potential use in anticancer therapy (40). The role that macrophages serve in carcinogenesis has been the focus of several studies, including systematic reviews (41-43).
M1 macrophages promote tumor elimination, whereas M2 macrophages facilitate carcinogenesis (44). As demonstrated over half a century ago, M1 macrophages are capable of killing and eliminating cancer cells in accordance with their primary physiological function, the elimination of foreign and harmful substances (45).
M1 cells initiate cytokine production in the tumor microenvironment and facilitate cancer cell destruction by recruiting pro-immunostimulatory leukocytes and phagocytizing tumor cells (46,47). M2 macrophages serve a leading role in tumor spread (48). M2 macrophages have a notable effect on tumor development in both the primary and metastatic foci. Their effects are associated with basement membrane degradation, angiogenesis and general immunosuppression (49,50).
Macrophages have been shown to be present not only in the M1 or M2 states in the tumor microenvironment, but also in transitional states, and the role of the transitional states in tumorigenesis remains poorly understood (51). The elimination of all macrophage populations regardless of the polarization state may provide a potentially effective approach to therapy as both primary and metastatic tumorigenesis is reduced as a result (52).
The activation of macrophages is widely regarded as polarization in the direction of the M1 or M2 states. However, the M2 activation state includes heterogeneous and functionally distinct macrophages. Studies on the existence of macrophages of the M2a, M2b, M2c and M2d phenotypes make it possible to specify a number of aspects regarding the nature of the immune response (Table I).
Table I
Classification of phenotypes of M2 macrophages.
| Phenotype of M2 macrophages | Stimulus for polarization | Produced cytokines | Functions |
|---|---|---|---|
| М2а | IL-4, IL-13 | IL-10, TGF-β, IL-1 receptor antagonist | Activation of Th2-reactions, eosinophil involvement, connective tissue growth |
| М2b | IL-1β, immunocomplex + lipopolysaccharide | IL-1, IL-6, IL-10, TNF-α | Regulation of inflammatory and immune responses, activation of Th2 |
| М2с | IL-10, TGF-β, glucocorticosteroids | IL-10, TGF-β | Remodeling, intercellular matrix synthesis |
| М2d | IL-6, adenosine | IL-10, IL-12, TNF-α, TGF-β | Tumor metastasis and progression |
[i] IL, interleukin; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-α.Table I – Classification of phenotypes of M2 macrophages.
M2a and M2b phenotype macrophages typically exhibit anti-inflammatory activity. Macrophages of the M2c phenotype are very similar to M1 macrophages, with the exception of high (increased) IL-10 expression as opposed to pro-inflammatory cytokines (53).
Wang et al (54) isolated the M2d phenotype, characterized by decreased secretion of IL-12 and increased secretion of IL-10. M2d macrophages are common in the tumor microenvironment. It is hypothesized that M2d macrophages are induced following stimulation with Toll-like receptor agonists and adenosine, and/or tumor-related factors. Isolation of subtypes of macrophages of the M2 family may facilitate the possibility of their targeted therapeutic use for treatment of tumors.
However, Quail and Joyce (55) demonstrated that the clinical effect of isolation of subtypes of macrophages of the M2 family was limited due to the limitations in the methods of targeted drug delivery to macrophages. However, macrophages preserve their plasticity regardless of polarization and, in particular, remain capable of switching from one phenotype to another dependent on the stimuli from the microenvironment (55).
The presence of macrophages in primary tumors is associated with a poor prognosis (56-59), with colorectal cancer serving as the only exception (60). M1 and M2 macrophages present in the tumor microenvironment have been the focus of an increasing number of studies (20,28,51).
Although the causal associations have not yet been established, the available body of research highlight the possibility of novel therapeutic strategies that are aimed at eliminating macrophages or altering the macrophage phenotype (61).
Since increased TAM infiltration is associated with a poor prognosis and therapeutic failure in cancer, TAM reprogramming toward the anticancer M1 phenotype and TAM suppression may provide promising strategies for the treatment of cancer (62).
Prospective use of TAMs for anticancer therapy
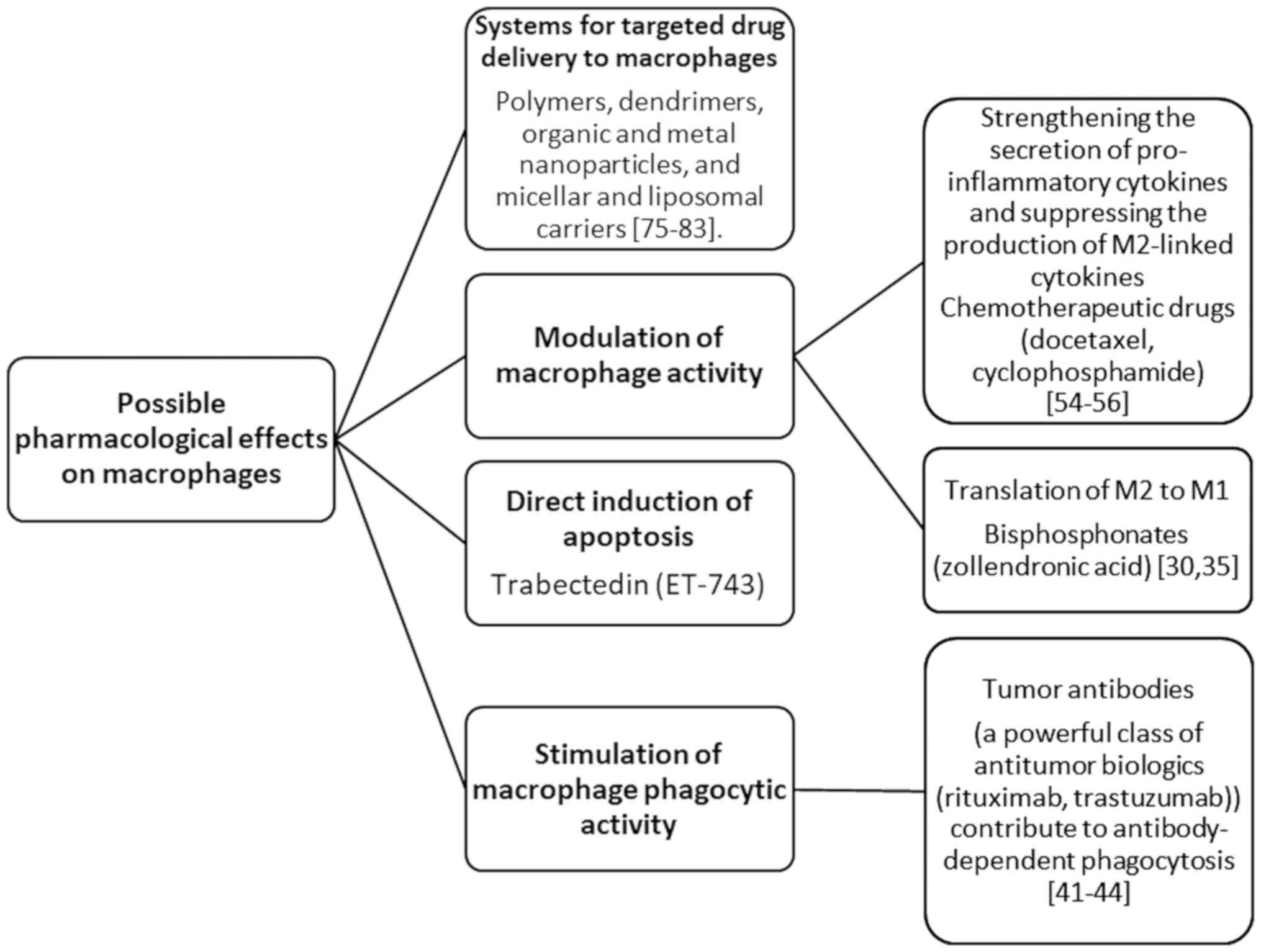
Based on the literature search performed for the present review, three macrophage-related strategies of cancer therapy are speculated. These strategies involve drugs that modulate TAM activity; engineered carriers for targeted drug delivery to macrophages, TAMs, or specific pro-tumoral M2-TAMs; and macrophage self-targeting to the tumor.
Drugs modulating macrophage activity
Various drugs that modulate macrophage activity are illustrated in Fig. 2.
Bisphosphonates modulate macrophage activity and are used in the treatment of bone tissue disorders, such as osteoporosis and bone metastases in cancer. A previous preclinical study using a mouse model of breast tumors suggested that an extra skeletal therapeutic effect is additionally exerted by bisphosphonates (63).
Zoledronic acid, which is a medication used in the treatment of cancer, has been shown to revert macrophage polarization from the M2 to the M1 phenotype, thus inhibiting spontaneous breast carcinogenesis (64). Zoledronic acid acts as a potent inhibitor of farnesyl pyrophosphate synthase, which is a key enzyme of the mevalonate pathway.
By inhibiting farnesyl pyrophosphate synthase, zoledronic acid prevents the prenylation of small G-proteins, such as Ras, Rho and Rap1A, which are necessary for cancer cell adhesion, migration and invasion. It has been shown that zoledronic acid binds primarily with microcalcifications present in breast tumors and is then phagocytized by TAMs, leading to apoptosis and M2-to-M1 transformation. It has been demonstrated in vivo that zoledronic acid inhibits the production of the proangiogenic factor, matrix metalloproteinase, and triggers the TAM transition from the pro-tumoral M2 phenotype to the antitumor M1 phenotype (65).
In 2013, Rogers (66) examined the antitumor effects of zoledronic acid in vitro and in vivo by evaluating its effect on macrophages. J774 macrophages were treated with zoledronic acid alone and in combination with doxorubicin in vitro and apoptosis and necrosis were evaluated.
Following treatment with zoledronic acid, its uptake was estimated by detecting unprenylated Rap1A (uRap1A) in J774 macrophages in vitro, and in peritoneal macrophages and macrophage populations from subcutaneous breast cancer xenografts in vivo.
The treatment of J774 macrophages with 5 µM zoledronic acid for 24 h significantly increased the uRap1A levels, while apoptotic cell death was induced at higher concentrations or longer exposure times. Doxorubicin (10 nM, 24 h) and zoledronic acid (10 µM, 24 h) used consecutively increased cell death compared with that observed with the use of either drug alone.
Detectable uRap1A levels were observed in peritoneal macrophages and macrophage populations isolated from breast tumor xenografts 24 h after single administration of zoledronic acid at 100 µg/kg in vivo (66). Zoledronic acid concentrations <10 nM were shown to inhibit Rab prenylation in J774 macrophages following long-term exposure in culture. Quantitative mass spectrometry identified 18 different unprenylated Rab proteins and revealed that their accumulation increased at least 7-fold following the treatment of J774 cells with nanomolar concentrations of zoledronic acid (67).
Another study demonstrated that zoledronic acid combined with ultrasonic treatment was significantly more effective than zoledronic acid alone (P<0.01). The B02 tumor size in mice treated with zoledronic acid and ultrasound was 42% lower (P<0.002) compared with mice treated with bisphosphonate alone (68). Bisphosphonates are administered in liposomes or attached to nanoparticles to improve their pharmacokinetics, reduce the side-effects and to alter their biodistribution (65). Liposomal bisphosphonate forms are capable of inducing the M2-to-M1 phenotypic transition (69).
Thus, studies on bisphosphonates used alone or in combination with anticancer drugs or physicochemical methods for the treatment of tumorigenesis are promising fields of research, and highlight possibility of developing novel therapeutic strategies (70-74).
Drug-dependent stimulation of phagocytic activity to modulate macrophages
The phagocytosis of foreign bodies, apoptotic cells and cancer cells, and the stimulation of adaptive immunity by presenting the antigens of assimilated materials, are two important innate immune functions of macrophages (75). Tumor-specific antibodies are a class of potent biopharmaceuticals, which act by directly inhibiting the transmission of survival signals, mediating antibody-dependent cell cytotoxicity of natural killer cells, inducing complement-dependent cytotoxicity via the activation of the complement cascade, and thus promoting antibody-dependent cell phagocytosis by macrophages (76). Studies have indicated that monoclonal antibodies approved as anticancer drugs, such as rituximab and trastuzumab, exert their therapeutic effects mostly through antibody-dependent cell phagocytosis (77,78). In spite of their potential to stimulate tumor cell invasion, TAMs are capable of tumor cell phagocytosis in the presence of target antibodies (79,80).
Thus, to improve the therapeutic strategy based on stimulating antibody-dependent cell phagocytosis, the Fc fragments of antibodies should be engineered to increase their interaction with receptors on macrophages (81). Although IgG class antibodies are typically used to design antibody-based therapeutics, the therapeutic potential of other antibody isotypes (IgA and IgE) has been the subject of several preclinical studies, where monocytes/macrophages also serve an important role in affecting the functions of antibody-dependent cell cytotoxicity and antibody-dependent cell phagocytosis (82,83).
Chemotherapeutic drugs are also considered potential means with which to modulate macrophages. A number of anticancer chemotherapeutics exert their pharmacological effects on non-tumor cell populations, although additional studies are required in the field, as the current literature is limited to preliminary results from in vitro experiments (84-89) In particular, trabectedin and lurbinectedin (a second-generation analog) are efficient in eliminating TAMs (84,85). Trabectedin mechanically interacts with the TRAIL-R2 ligand-receptor, and induces tumor necrosis factor-related apoptosis of mononuclear phagocytes by causing receptor clustering and subsequent caspase 8-dependent apoptosis activation (86).
In addition to exerting cytotoxic effects, certain chemotherapeutics modulate the macrophage response to the tumor (15,87). A previous study using mouse models of fibrosarcoma and breast tumors demonstrated that docetaxel promotes target cell polarization to macrophages with an antitumor M1 phenotype (88).
Cyclophosphamide treatment facilitates macrophage infiltration, increases the secretion of proinflammatory cytokines (IL-6 and IL-12) and suppresses the production of pro-tumoral M2-associated cytokines (IL-4, IL-10 and IL-13) (89,90). As a mechanism of self-protection against chemotherapy, chemoresistant cancer cells secrete IL-34, which increases their survival and promotes the polarization of TAMs towards an M2 phenotype to further facilitate an immunosuppressive environment (91). Thus, a combination of chemotherapy and immunotherapy may be more efficient in inducing tumor regression.
reference link: https://www.spandidos-publications.com/10.3892/br.2020.1354
More information: Valentí Gómez et al. Breast cancer–associated macrophages promote tumorigenesis by suppressing succinate dehydrogenase in tumor cells, Science Signaling (2020). DOI: 10.1126/scisignal.aax4585



{kind=link}