









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



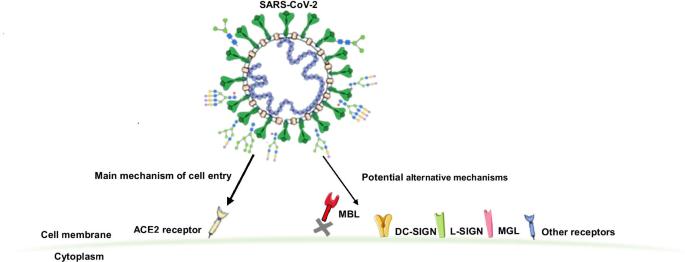
Un nuovo studio condotto da ricercatori del Genos Glycoscience Research Laboratory-Croazia e dell’Università di Zagabria-Croazia ha sorprendentemente scoperto che il coronavirus SARS-CoV-2 è in grado non solo di legarsi ai recettori ACE2 , ma è anche in grado di legarsi ad altri 13 recettori dell’ospite umano .
Questi recettori dell’ospite includono:
1. C-type lectin receptors
2. Mannose-binding lectin (MBL)
3. DC-SIGN including
4. L-SIGN
5. Macrophage galactose type C-type lectin (MGL)
6. Mannose receptor
7. Lectin-like oxidized low-density lipoprotein receptor-1
8. Liver and lymph node sinusoidal endothelial cell C-type lectin
9. Toll-like receptors (TLRs)
10. Neuropilin-1
11. Glucose-regulated protein 78 (GRP78)
12. Siglecs
13. Galectins
Alcuni di questi recettori sono stati identificati come potenziali recettori nei primi studi ma non sono mai stati seguiti.
Alcuni altri scienziati che non sono stati coinvolti nello studio ipotizzano che, ancora di più, l’ emergente SARS-CoV-2 e le varianti con mutazioni più recenti sulle loro proteine spike si stanno adattando in modo tale da essere in grado non solo di legarsi ad altri recettori dell’ospite umano, ma anche di eludere le risposte immunitarie umane o addirittura di interrompere e spegnere le risposte immunitarie umane per un po’.
Alcune delle possibili conseguenze di ciò è che vedremo più persone inizialmente infettate che esprimono manifestazioni sintomatiche asintomatiche o semplicemente lievi mentre il SARS-CoV-2 potrebbe causare più danni alle vie cellulari, ai tessuti e agli organi umani attraverso nuove modalità di patogenesi , con conseguente condizioni mediche più croniche a lungo termine!
Questo è qualcosa che deve essere studiato di più mentre il mondo è in una fase di apertura e allentamento delle restrizioni sul COVID-19 e molti presumendo che la malattia diventerà endemica e tutti stanno prendendo le cose alla leggera mentre il mondo potrebbe avere una catastrofica crisi sanitaria incombente con Long COVID o PASC.
Secondo il team di studio, “L’infezione da coronavirus SARS-CoV-2 mostra una vasta gamma di manifestazioni cliniche. Sebbene siano stati identificati alcuni fattori di rischio per la gravità e gli esiti di COVID-19, i meccanismi biologici sottostanti non sono ancora ben compresi.Le proteine SARS-CoV-2 di superficie sono fortemente glicosilate consentendo l’interazione della cellula ospite e l’ingresso virale.
L’enzima di conversione dell’angiotensina 2 (ACE2) è stato identificato come il principale recettore della cellula ospite che consente l’ingresso delle cellule SARS-CoV-2 dopo l’interazione con la sua glicoproteina S.
Tuttavia, studi recenti riportano l’interazione della glicoproteina SARS-CoV-2 S con altri recettori cellulari, principalmente lectine di tipo C che riconoscono epitopi glicani specifici che facilitano l’ingresso di SARS-CoV-2 nelle cellule sensibili. “
I risultati dello studio hanno rivelato che SARS-CoV-2 è stato in grado di interagire e legarsi ad altri recettori di superficie della membrana cellulare e lectine solubili, influendo sull’ingresso delle cellule virali, modulandone l’infettività e potenzialmente svolgendo un ruolo nelle successive manifestazioni cliniche di COVID-19.
I risultati dello studio sono stati pubblicati sulla rivista peer review: Glycoconjugate Journal di Springer. https://link.springer.com/article/10.1007/s10719-021-10021-z
La proteina Spike (S) del coronavirus sSARS-CoV-2 è parzialmente responsabile della patogenicità del virus. È ben noto che il dominio di legame al recettore (RBD) della subunità S1 della proteina S può legarsi ai recettori dell’enzima di conversione dell’angiotensina 2 (ACE2), consentendo così l’ingresso del virus nella cellula ospite. Comparativamente, il dominio terminale N della subunità S2 è responsabile della fusione della membrana.
Tuttavia, rapporti recenti hanno dimostrato che la glicoproteina S è in grado di interagire con più recettori cellulari alternativi.
Il team di studio del laboratorio di ricerca Genos Glycoscience in Croazia ha recentemente studiato i siti con cui la proteina S è in grado di legarsi.
È noto che ogni protomero della glicoproteina S contiene 22 siti di N-glicosilazione e tre siti di O-glicosilazione.
Gli studi precedenti, tuttavia, mostrano risultati incoerenti nella caratterizzazione della glicosilazione, con alcuni ricercatori che mostrano la N-glicosilazione completa per la maggior parte del tempo, e altri solo una glicosilazione parziale.
https://academic.oup.com/glycob/article/30/12/981/5826952
https://www.biorxiv.org/content/10.1101/2020.07.29.227462v1
Inoltre c’è stato anche disaccordo sul tipo di glicani trovato, con studi alternativi che mostrano ibridi, alto mannosio e N-glicani complessi.
Questa ampia variazione può essere dovuta al metodo con cui la glicoproteina S è glicosilata, poiché utilizza i meccanismi di glicosilazione degli ospiti che alla fine si traducono in modelli simili alle cellule ospiti. Ciò comporterebbe diversi modelli di glicosilazione e diversi tipi di glicani, a seconda del tipo di cellula in cui è stato costruito il particolare virus.
È anche noto che il legame del RBD SARS-CoV-2 ad ACE2 è facilitato dall’interazione glicosaminoglicano eparan solfato del glicocalice cellulare, che aiuta la struttura della proteina S a una conformazione più aperta. Tuttavia, mentre ACE2 è il recettore principale per l’ingresso delle cellule SARS-CoV-2, mostra una bassa espressione nel sistema respiratorio.
https://pubmed.ncbi.nlm.nih.gov/32970989/
https://www.embopress.org/doi/full/10.15252/msb.20209610
https://jamanetwork.com/journals/jama/fullarticle/2766524
https://www.biorxiv.org/content/10.1101/2020.05.29.123513v1
Considerando che SARS-CoV-2 si diffonde principalmente tra gli ospiti attraverso l’infezione da goccioline, ciò indica che la glicoproteina S può interagire con altri recettori per entrare nella cellula .
La potente spettroscopia di risonanza magnetica nucleare (NMR) ha rivelato che gli N-glicani del RBD possono legarsi a varie lectine, tra cui la lectina del galattosio dei macrofagi (MGL), le galectine -3, -7 e -8, nonché l’immunoglobulina legante l’acido sialico lectina.
https://onlinelibrary.wiley.com/doi/10.1002/anie.202011015
È interessante notare che i livelli di espressione di MGL sono risultati elevati nei pazienti con grave malattia da coronavirus 2019 (COVID-19). Questa osservazione suggerisce potenzialmente che il virus potrebbe rispondere alle reazioni immunitarie attraverso l’ingresso delle cellule attraverso questi recettori.
Potenzialmente più allarmante è la possibilità che la glicoproteina S si leghi ai recettori della lectina di tipo C (CLR). Questi possono essere legati da N-glicani oligomannosi; infatti, circa 1 su 3 glicani SARS-CoV-2 S sono di questo tipo.
I CLR si trovano principalmente sulle cellule che presentano l’antigene, come i macrofagi e le cellule dendritiche. Entrambi questi tipi di cellule si trovano spesso nei polmoni.
Infatti, quando si verifica un’infiammazione, anche i monociti che circolano nel sangue e in altri tessuti possono differenziarsi in queste cellule. Ciò fornirebbe sia una via iniziale per l’infezione nel sistema respiratorio se ACE2 mostrasse una bassa espressione, sia per l’ingresso virale nelle cellule del flusso sanguigno.
Inoltre, la lectina galattosio macrofagica (MGL) è un altro recettore espresso principalmente nelle cellule dendritiche e nei macrofagi nei polmoni umani e nel sistema respiratorio. MGL riconosce i glicani che portano il galattosio terminale.
Gli studi trattati in varie notizie COVID-19 hanno già dimostrato che SARS-CoV-2 è in grado di legarsi a questa lectina, supportando ulteriormente la convinzione che il virus abbia vie alternative per l’ingresso nelle cellule.
Un ulteriore esame del sequenziamento degli amminoacidi suggerisce che entrambi i glicani N e O sono coinvolti nel legame con MGL. https://pubmed.ncbi.nlm.nih.gov/34015061/
Il team di studio ha scoperto che gli studi esaminati mostrano che la glicoproteina SARS-CoV-2 S può legarsi a più altri recettori, entrambi consentendo la transinfezione di cellule sensibili, oltre a consentire l’infezione di più tipi di cellule.
La transinfezione delle cellule non solo porterà a una maggiore trasmissione virale, ma poiché i tipi di cellule vulnerabili alla transinfezione sono principalmente cellule immunitarie, ciò potrebbe esacerbare il rischio di grave infiammazione e rilascio di citochine.
Inoltre, le interazioni mediate dalla glicosilazione con SARS-CoV-2 potrebbero suggerire che la glicosilazione di SARS-CoV-2 potrebbe avere effetti significativi sulla gravità della malattia nel paziente, nella misura in cui potrebbe determinare il recupero o la diffusione.
Ciò suggerisce un meccanismo per l’infezione asintotica da SARS-CoV-2, poiché una diminuzione del titolo di SARS-CoV-2 dovuta all’endocitosi da parte delle cellule immunitarie nei polmoni può impedire la comparsa dei sintomi.
Ulteriori ricerche urgenti e dettagliate sono giustificate poiché le implicazioni dei risultati dello studio sono preoccupanti sotto molti aspetti.
Un dominio di legame al recettore (RBD) è una parte fondamentale di un virus situato sul suo dominio “spike” che gli consente di agganciarsi ai recettori del corpo per entrare nelle cellule e portare all’infezione.
Questi sono anche gli obiettivi primari nella prevenzione e nel trattamento delle infezioni virali, inclusa la sindrome respiratoria acuta grave coronavirus 2 (SARS-CoV-2), il virus che causa il COVID-19.
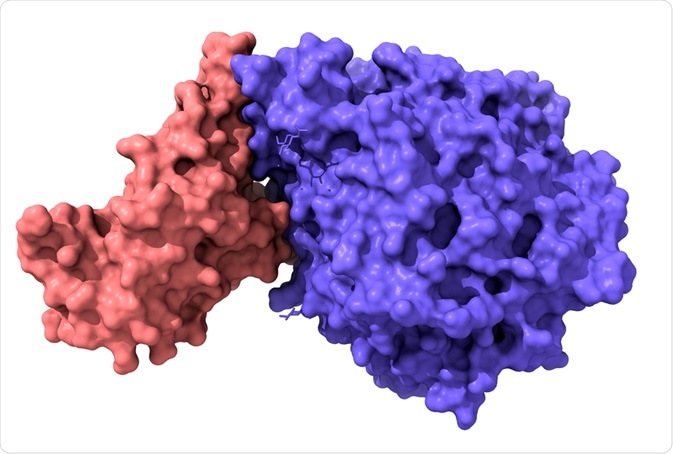
Che cos’è un dominio di legame al recettore (RBD)?
Un dominio di legame al recettore (RBD) è un breve frammento immunogenico di un virus che si lega a una specifica sequenza di recettori endogeni per ottenere l’ingresso nelle cellule ospiti. In particolare, questi si riferiscono a una parte della glicoproteina “spike” (dominio S) che è necessaria per interagire con i recettori endogeni per facilitare la fusione della membrana e la consegna al citoplasma. Tipicamente, il dominio S è anche il sito di anticorpi neutralizzanti.
SARS e MERS
L’ingresso di beta-coronavirus, come il coronavirus della sindrome respiratoria acuta grave (SARS-CoV), il virus che causa la SARS, richiede il legame della sua glicoproteina spike, il dominio “S”, al recettore ACE2 nel corpo. I coronavirus sono punteggiati da tali domini S su tutta la loro superficie dando l’aspetto di un grande aspetto distintivo di “corona” e quindi il nome “corona”/corona, virus.
Ogni dominio S è una proiezione superficiale di circa 20 nm che circonda la periferia del coronavirus e varia considerevolmente tra i diversi coronavirus. All’interno del dominio S di SARS-CoV, c’è un breve dominio (all’interno della subunità S1) contenente solo 2 siti di glicosilazione che secernono brevi frammenti di RBD (per glicosilazione) che si piegano e si legano al recettore ACE2.
Un sito di glicosilazione funzionale è sufficiente per creare un frammento RBD che può legarsi ad ACE2. Mutazioni specifiche all’interno dei residui di amminoacidi RBD (ad es. K390) portano a una diminuzione dell’affinità per ACE2. Pertanto, il legame dell’RBD virale sul suo dominio S è un passaggio essenziale per la fusione della membrana e l’ingresso nelle cellule ospiti bersaglio, poiché porta il dominio S2 a transitare da uno stato di prefusione a uno stato di postfusione stabile ancoraggio lo alla membrana.
Per quanto riguarda il coronavirus correlato alla sindrome respiratoria mediorientale (MERS-CoV), il virus che causa il MERS, la differenza chiave tra MERS-Cov e SARS-CoV è il dominio S virale (S1). In MERS-CoV, una glicoproteina di membrana di tipo I da 1353 aminoacidi si assembla in trimeri formando i “picchi” che si legano e si fondono al recettore DPP4/CD26.
Il MERS-CoV RBD (E367 e Y606) forma un complesso con il dominio extracellulare di DDP-4 (S39-P766). Pertanto, mentre i domini CoV principali rimangono molto simili, la loro sequenza RBD varia considerevolmente, influenzando la diversa affinità del recettore.
SARS-CoV-2 (COVID-19)
Simile a SARS-CoV, SARS-CoV-2 (il virus che causa COVID-19) si lega al recettore ACE2 per entrare nelle cellule epiteliali respiratorie e digestive. Il dominio S di SARS-CoV-2 contiene un RBD che gli consente di legarsi ad ACE2 e fondersi nella membrana delle cellule epiteliali.
L’RBD del dominio S1 di SARS-CoV-2 si lega fortemente all’ACE2 sia umano che di pipistrello. L’RBD di SARS-CoV-2 si trova tra i residui 331-524 (o Thr333-Gly526 in uno studio diverso) del dominio S1, che gli consente di legarsi ad ACE2 – e più fortemente di SARS-CoV (secondo alcuni studi) – che può riflettere la maggiore infettività di SARS-CoV-2 rispetto a SARS-CoV.
Il SARS-CoV-2 RBD ha un foglietto beta antiparallelo a 5 filamenti intrecciati con brevi eliche e anelli di collegamento. Nel nucleo, tra i fili β4-7, c’è un’ulteriore inserzione estesa contenente brevi fili β5-6. Questa estensione è dove si trova il motivo di legame al recettore (RBM), che contiene i residui di contatto che gli consentono di legarsi all’ACE2.
L’identificazione del RBD può aiutare a portare allo sviluppo di anticorpi monoclonali (o policlonali) (vaccini) nel trattamento e nella prevenzione dell’infezione coronavirale, poiché il RBD è il sito per molti dei principali anticorpi neutralizzanti, impedendo il legame del virus al recettore ( ACE2/DPP4).
Gli studi hanno dimostrato che gli anticorpi policlonali specifici per SARS-CoV RBD possono cross-reagire con la proteina RBD SARS-CoV-2 per inibirne l’ingresso in ACE2 in vitro (cellule che esprimono umani). Inoltre, gli anticorpi policlonali SARS-CoV RBD possono anche neutralizzare in modo incrociato l’infezione pseudovirale SARS-CoV-2 portando alla possibilità di creare un vaccino basato su SARS-CoV-RBD per la prevenzione di SARS-CoV-2.
Pertanto, il plasma convalescente di pazienti guariti da SARS e COVID-19 viene esaminato in termini di trattamento e prevenzione per COVID-19. Molti studi sono in corso sul plasma convalescente in COVID-19 e suggeriscono che non è efficace in seguito nella malattia grave e ospedalizzata, ma potrebbe ancora essere un’opzione per il trattamento precoce o preventivo.
Molti anticorpi raccolti contro SARS-CoV hanno mostrato risultati contrastanti riguardo a SARS-CoV-2, con un solo (CR3002) che può legarsi a entrambi che mira a una porzione di SARS-CoV-2 RBD.
Il plasma convalescente da pazienti con SARS guariti che ha una reazione crociata con SARS-CoV-2 richiede un’analisi più approfondita della sequenza dell’RBD che è interessata nello sviluppo di vaccinazioni preventive forti e di successo. Pertanto, lo studio del RBD di SARS-CoV-2 è importante per terapie e vaccinazioni di successo. Molte diverse vaccinazioni sono state ora create e somministrate con successo per SARS-CoV-2.
Riferimenti
- Chakraborti et al, 2005. The SARS Coronavirus S Glycoprotein Receptor Binding Domain: Fine Mapping and Functional Characterization. Virology Journal 2(73). https://virologyj.biomedcentral.com/articles/10.1186/1743-422X-2-73
- He et al, 2004. Receptor-binding domain of SARS-CoV spike protein induces highly potent neutralizing antibodies: implication for developing subunit vaccine. Biochem Biophys Res Commun. 324(2): 773–781. https://pubmed.ncbi.nlm.nih.gov/15474494/
- Mou et al, 2013. The Receptor Binding Domain of the New Middle East Respiratory Syndrome Coronavirus Maps to a 231-Residue Region in the Spike Protein That Efficiently Elicits Neutralizing Antibodies. J Virol. 87(16): 9379–9383. https://pubmed.ncbi.nlm.nih.gov/23785207/
- Tai et al, 2020. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: implication for the development of RBD protein as a viral attachment inhibitor and vaccine. Cell Mol Immunol. 17(6): 613–62. https://pubmed.ncbi.nlm.nih.gov/32203189/
- Lan et al, 2020. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 581(7807):215-220. https://pubmed.ncbi.nlm.nih.gov/32225176/
- Premkumar et al, 2020. The receptor-binding domain of the viral spike protein is an immunodominant and highly specific target of antibodies in SARS-CoV-2 patients. Sci Immunol. 5(48):eabc8413 https://pubmed.ncbi.nlm.nih.gov/32527802/



{kind=link}