










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new study by researchers from Institut Pasteur-France has found that SARS-CoV-2 uses tunneling nanotubes (TNT) to spread between permissive cells and non-permissive neuronal cells.
To date the mechanism through which SARS-CoV-2 infects neurons is not understood.
However, it has been known that Tunneling nanotubes (TNTs) ie actin-based intercellular conduits that connect distant cells, allow the transfer of cargos, including viruses.
The study team explored the neuroinvasive potential of SARS-CoV-2 and whether TNTs are involved in its spreading between cells in vitro.
The study findings show that neuronal cells, not permissive to SARS-CoV-2 through an exocytosis/endocytosis dependent pathway, can be infected when co-cultured with permissive infected epithelial cells. SARS-CoV-2 induces TNTs formation between permissive cells and exploits this route to spread to uninfected permissive cells in co-culture.
The study findings were published in a preprint server and are currently being peer reviewed. https://www.biorxiv.org/content/10.1101/2021.11.15.468633v1
However, how SARS-CoV-2 gains access to the CNS and how infection leads to neurological symptoms is still not clear (14–18). SARS-CoV-2 neuroinvasion could be achieved through several routes as previously described (19), and once it reaches the CNS it could bind the angiotensin-converting enzyme 2 (ACE2) receptor exposed on neuronal cells to infect the brain (20).
ACE2 receptor is the main actor responsible of the virus entry in the lower respiratory tract (13,21,22). To enter host cells the viral spike (S) proteins of coronaviruses bind the enzymatic domain of the ACE2 receptor, which is exposed on the surface of the cells forming the oral cavity and the oropharynx (23–25).
While the expression of the ACE2 receptor has been well documented in many cell type and tissues (23, 24), it is important to underline that in human brain the expression of the ACE2 receptor is low, with the exception of brain areas such as the thalamus and the choroid plexus (26).
For this reason, it is not clear how the virus can propagate through the brain, and it is a priority to investigate how SARS-CoV-2 enters into neuronal cells in order to provide new insights in the understanding of the neurological manifestations associated with COVID-19.
An interesting aspect to consider is that the presence of SARS-CoV-2 RNA and proteins has been found in anatomically distinct regions of the brain of COVID-19 patients (13, 27). In this respect, the spreading of SARS-CoV-2 in the CNS is reminiscent of toxic amyloid proteins in neurodegenerative disorders (NDs), that propagate in the brain according to the progression of the pathology (28, 29).
We have previously shown that the spreading of different amyloid aggregates between cells of the CNS and from peripheral lymphoid system cells to neurons occurs mainly through Tunneling Nanotubes (TNTs), a novel mechanism of cell-to-cell communication (30–32). TNTs are thin, membranous conduits rich in actin that form contiguous cytoplasmic bridges between cells over long and short distances (33, 34).
Recently we set up a pipeline in correlative cryo-electron microscopy (Cryo-CLEM) to determine the ultrastructure of TNTs in neuronal cell lines, demonstrating their structural identity and differentiating them from other cellular protrusion as filopodia (35). Functionally, TNTs allow direct transport of cargos including virus between distant cells (30,34,36,37).
Of specific interest, it has been documented that viruses from different families induce increased formation of TNTs or TNT-like structures, using these membranous structures to efficiently spread the infection to neighboring health cells (38–42).
Here, we investigated the neuroinvasive potential of SARS-CoV-2 and whether TNTs are involved in its intercellular spreading. Since TNT-transferred virions would not necessarily be exposed outside the host cell, we hypothesize that TNT-mediated transmission can be used by the viruses to escape neutralizing antibody activity and immune surveillance, as well as to infect less permissive cells lacking the membrane receptor for virus entry, thus allowing for spreading of virus tropism and pathogenicity.
We observed, in confocal microscopy, that SARS-CoV-2, induced the formation of TNTs that then could be used by the virus to efficiently spread toward uninfected permissive and non-permissive cells.
Furthermore, by setting up correlative Cryo-CLEM and–tomography (35), we demonstrated that SARS-CoV-2 virions are associated to the plasma membrane of TNTs formed between permissive cells. Interestingly, we also observed virus-like vesicular structures and double membrane vesicles (DMVs) inside the TNTs, both between permissive cells and between permissive and non-permissive cells.
Altogether, our results shed new light on the structure of the viral particles undergoing intercellular spreading and provide important information about the molecular mechanism of SARS-CoV-2 infection and transmission. They support the role of TNTs in the viral spreading in both permissive and non-permissive cells, possibly enhancing the efficiency of viral propagation through the body.
. . . . .
In a past study, researchers have shown a novel mechanism of cell-to-cell communication by spreading different amyloid aggregates between the CNS and from the peripheral lymphoid system cells to neurons via TNTs.
https://pubmed.ncbi.nlm.nih.gov/19198598/<;/a>
https://pubmed.ncbi.nlm.nih.gov/27550960/
https://pubmed.ncbi.nlm.nih.gov/31625188/
Tunneling nanotubes or TNTs are dynamic connections between cells that consist of thin membranous conduits rich in actin that form contiguous cytoplasmic bridges between cells over long and short distances. These TNTs are capable of transporting cargo, including viruses, between cells.
Numerous studies have also reported the formation of TNTs or TNT-like structures that have been induced by HIV virus, retroviruses, herpesviruses, influenza A, and the human metapneumovirus to enable the efficient spread of infection to neighboring healthy cells. https://pubmed.ncbi.nlm.nih.gov/18835599/
https://pubmed.ncbi.nlm.nih.gov/32024778/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3232570/
https://pubmed.ncbi.nlm.nih.gov/30917314/
https://pubmed.ncbi.nlm.nih.gov/18193035/
Role of Tunneling Nanotubes in Viral Infection, Neurodegenerative Disease, and Cancer
The ability of a cell to communicate with the distant neighboring cell is vital for its survival and efficient function. Many pharmaceuticals work by enhancing or inhibiting components of the cell communication mechanisms (1, 2). The formation of filamentous (F)-actin rich tubules termed tunneling nanotubes (TNTs) are one such mechanism of communication between the cells (3, 4).
TNTs have been broadly defined as thin membrane tubes which connect two cells and mediate the transfer of cellular cargo (5, 6). A recent review has added more detail to this definition, stating that TNTs must satisfy three requirements: they must connect two or more cells, be composed of F-actin, and not come in contact with the substrates that pass through them (7). In this review, we use the more specific definition to differentiate TNTs from similar but distinct structures.
TNTs exist as a cytoplasmic bridge between the two closely or distant cells. They form gap-like junctions between connected cells and mediate the exchange of cytoplasmic proteins, cellular organelles (such as endoplasmic reticulum, Golgi, endosome, lysosome, mitochondria), lipids, nucleic acids, microRNA, ions, calcium, and several other components (4, 8–10).
The diameter of the TNT restricts the type of cargo that can be transported. Thinner TNTs contain F-actin and are less than 7 µm in diameter which prevents them from transferring large organelles between cells (11). Thicker TNTs contain microtubules and exceed a 7 µm diameter which allows them to transfer organelles such as mitochondria between cells (6, 12).
They facilitate both short and long-distance direct communication, spanning distances of up to 300 µm (13). TNTs are able to polymerize and depolymerize rapidly in 30-60 seconds, making them fluid, transient structures (8, 14). TNTs have been shown to be formed by different cell types including epithelial and fibroblasts (15–17), neuronal (18, 19), and multiple types of immune cells (20).
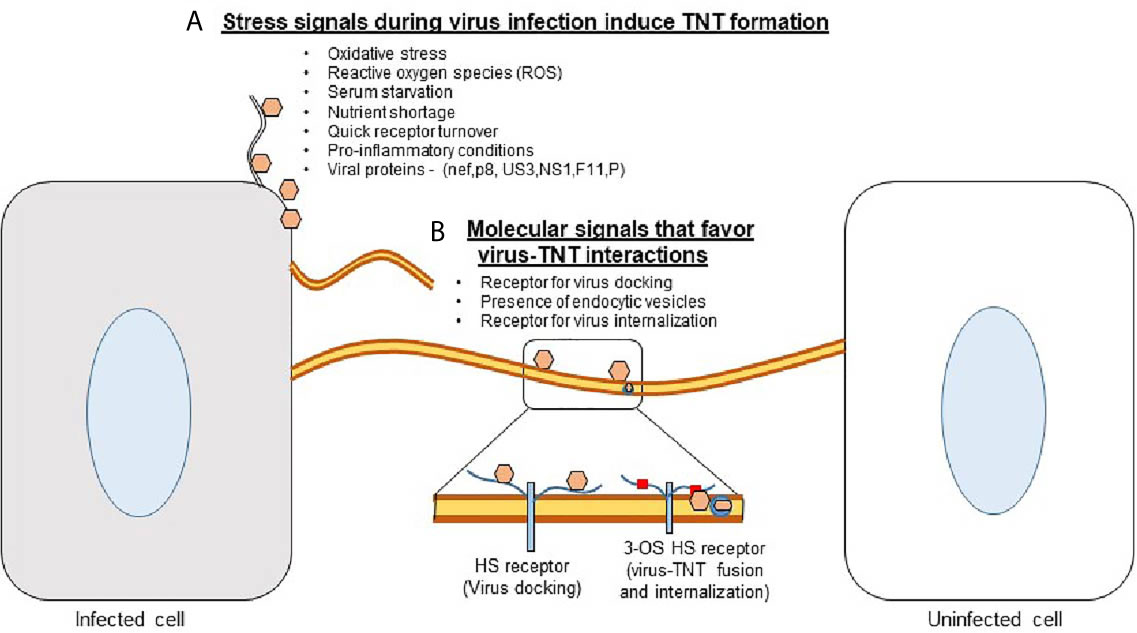
While numerous processes are downregulated when cellular supplies are low and cells are under distress, TNT polymerization is enhanced (21). Cellular stress such as viral infections, damage by UV light, or hydrogen peroxide-mediated oxidation have all been shown to upregulate TNT formation (Figure 1) (22).
TNT formation has recently been shown to play a large role in the pathogenesis of many diseases (23). Viral particles, prions, fungal spores, organelles, and other molecules can be transported by TNTs (23, 24). The concomitant evolution of pathogens with the cell’s ability to produce TNTs is interesting from a cellular biology standpoint and highlights the importance of targeting this mechanism of propagation to treat certain diseases.
reference link : https://www.frontiersin.org/articles/10.3389/fimmu.2021.680891/full



{kind=link}