










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Combination of nucleic acid nanotechnology and cryo-EM gives unprecedented insights into the structures of large and small RNAs, enabling fundamental advances in RNA biology and drug design.
Cryo-Electron Microscopy (Cryo-EM)
Electron Microscopy (EM) has become an extremely popular method for the ultrastructural study of macromolecules, cells, and tissues. An aqueous biological sample is frozen rapidly and irradiated with a beam of electrons. A detector senses how the electrons are scattered, and a computer reconstructs the 3D-shape of the molecule.
Why do you need this technology ?
- You have small amount of materials.
- The material is hard to crystallize.
- You are looking into the molecules in atomic details.
- It is a large biomolecular complex (>150 kDa, up to 2000 Å).
- You want to observe the real “native state” structure.
The Workflow of Our Cryo-EM (Take protein as an example)
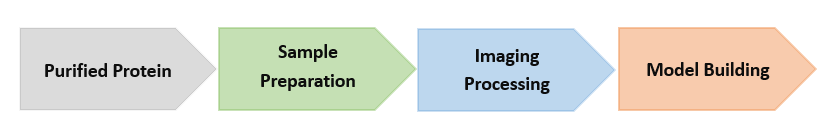
 | Sample preparation After protein purification, these samples will be treated with cryo-fixation. In this method, protein samples are placed on a specially treated EM grid consisting of tiny holes in a film supported by a metal frame. The grid is then plunged into liquid ethane to flash-freeze it, resulting in the protein samples being embedded in a thin layer of vitreous ice. Once the frozen-hydrated grid is prepared, it is placed in the electron microscope and kept at approximately -180 K throughout the experiment. |
 | EM imaging and data processingData collectionInitial 3D model calculationBeam-induced motion correctionMicrograph screeningAutomatic particle picking and normalization2D, 3D classification and refinement |
 | Model building and refinement Using our high-performance computers and software packages, we can interpret EM maps and reconstruct them into 3D structural models that satisfy the principles of physics and stereochemistry. (Our scientists are also very experienced with challenging targets such as membrane proteins by manual intervention to improve the initial fit or even build de novo models.) After an initial model is built, refinement is performed to maximize the agreement between the model and experimentally observed data by adjusting atomic coordinates, B factors, and other parameters. |
Single-particle electron cryo-microscopy (cryo-EM) has become the method of choice for the determination of protein structures. Cryo-EM analysis has several advantages over X-ray crystallography or NMR (1), but the method becomes increasingly challenging for smaller proteins. Large molecules are relatively easy to identify in noisy low-dose images of vitrified samples and have sufficient contrast and features to determine their orientation and position for alignment and averaging. The structural analysis of small particles (∼100 kDa or less) is much more difficult. Small targets often lack recognizable shape features that can facilitate initial image alignment at low resolution. Without symmetry, small particles require optimal conditions, such as a highly homogeneous sample, rigid protein conformation, and random particle distribution in thin ice, conditions that are difficult to achieve with most samples (2). However, structure determination of small proteins is of great interest, as most proteins have sizes below 100 kDa and ∼50% are smaller than 50 kDa, including many membrane proteins and proteins of medical importance. It is thus a major goal in the field to expand the use of cryo-EM to the routine analysis of small proteins.
One approach to employ cryo-EM for small proteins is based on phase contrast methods, such as the use of Volta phase plates. This method has been used to determine the structure of streptavidin, a protein of 52 kDa, at 3.2-Å resolution (3). However, the structure of this protein could be determined even without phase plates (4), likely because streptavidin forms rigid tetramers and the particles display a near-perfect distribution in very thin ice, which greatly facilitates structural analysis.
An alternative strategy is to make the target protein larger, either by fusing it to another protein or by using a binding partner. In either case, high rigidity of the added scaffold itself and its rigid connection to the target protein are required to facilitate particle alignment and averaging in cryo-EM images.
The fusion approach has been tried with different scaffolds. For example, in a recent study, the BRIL domain was fused into a loop of a small GPCR protein by extending helices on both sides of the fusion point; the size of the scaffold was further increased by a Fab directed against the BRIL domain (5). However, this approach is limited to proteins containing suitable α-helices; their extension has to be customized for each new target to generate a rigid connection, which is difficult to achieve without prior knowledge of the target structure.
More promising is the use of a binding partner that can be selected with a screening platform, such as modified ankyrin repeat proteins (DARPins), Fab fragments of antibodies, or nanobodies. In recent studies, DARPins selected against GFP were grafted onto large scaffolds and used to visualize GFP by cryo-EM (6, 7). However, the intrinsic conformational heterogeneity of DARPins limits their potential to achieve high-resolution structures of small proteins (7), and so far only a few DARPins have been selected against membrane proteins. Fab fragments can be used as a fiducial marker to facilitate image alignment in cryo-EM images (8), but they have been mainly used in X-ray crystallography. Only a few examples of their application for cryo-EM analysis have been reported (9–11), in part because the selection of appropriate Fabs is not trivial. In addition, the size of the Fabs (∼50 kDa) and the existence of a somewhat flexible hinge region between the two subdomains still make structural analysis challenging.
Nanobodies, derived from single-chain antibodies of camelids, are also becoming popular as versatile binding partners of target proteins. Nanobodies have several attractive features. They form rigid structures that can bind to diverse shapes of target proteins, such as loops, convex surfaces, and cavities (12). They can bind to small exposed surfaces, which may not be accessible to Fab fragments. Nanobodies can be selected from immunized camelids or from large in vitro libraries displayed by phages, yeast cells, or on ribosomes (12, 13), and can be produced in large quantities in a fairly short time. They often lock a protein into a fixed conformation, particularly in the case of membrane proteins, and have been used extensively to determine X-ray structures. The small size of nanobodies (∼12 to 15 kDa) limits their direct application in cryo-EM, but the problem might be overcome if one could increase their size with the rigid attachment of a large scaffold. One reported approach is to fuse a scaffold into a loop of the nanobody, generating a “megabody” (14). However, the linker consisted of β-strands between the nanobody and scaffold, which caused some flexibility and limited the use of the scaffold for particle alignment in cryo-EM analysis.
Here, we describe a versatile method that allows cryo-EM analysis of even the smallest protein once a tightly binding nanobody is available. The size of the nanobody is increased to ∼120 kDa by two rigidly attached scaffolds. The overall design is reminiscent of a Lego construction, so we propose to call the scaffolds/nanobody ensemble “Legobody.” The utility of the Legobody method is demonstrated by structures of two small proteins (22 kDa and 23 kDa) that are asymmetric monomers and have a size well below the estimated limit for direct cryo-EM single-particle analysis (∼40 kDa) (15). The Legobody approach can easily be applied to any target protein and should greatly expand the use of cryo-EM single-particle analysis by overcoming the current size limitations.
reference link : https://www.pnas.org/doi/full/10.1073/pnas.2115001118
We live in a world made and run by RNA, the equally important sibling of the genetic molecule DNA. In fact, evolutionary biologists hypothesize that RNA existed and self-replicated even before the appearance of DNA.
Fast forward to modern day humans: science has revealed that less than 3% of the human genome is transcribed into messenger RNA (mRNA) molecules that in turn are translated into proteins. In contrast, 82% of it is transcribed into RNA molecules with other functions many of which still remain enigmatic.
To understand what an individual RNA molecule does, its 3D structure needs to be deciphered at the level of its constituent atoms and molecular bonds. Researchers have routinely studied DNA and protein molecules by turning them into regularly packed crystals that can be examined with an X-ray beam (X-ray crystallography) or radio waves (nuclear magnetic resonance). However, these techniques cannot be applied to RNA molecules with nearly the same effectiveness because their molecular composition and structural flexibility prevent them from easily forming crystals.

Now, a research collaboration led by Wyss Core Faculty member Peng Yin, Ph.D. at the Wyss Institute for Biologically Inspired Engineering at Harvard University, and Maofu Liao, Ph.D. at Harvard Medical School (HMS), has reported a fundamentally new approach to the structural investigation of RNA molecules.
ROCK, as it is called, uses an RNA nanotechnological technique that allows it to assemble multiple identical RNA molecules into a highly organized structure, which significantly reduces the flexibility of individual RNA molecules and multiplies their molecular weight. Applied to well-known model RNAs with different sizes and functions as benchmarks, the team showed that their method enables the structural analysis of the contained RNA subunits with a technique known as cryo-electron microscopy (cryo-EM). Their advance is reported in Nature Methods.
“ROCK is breaking the current limits of RNA structural investigations and enables 3D structures of RNA molecules to be unlocked that are difficult or impossible to access with existing methods, and at near-atomic resolution,” said Yin, who together with Liao led the study. “We expect this advance to invigorate many areas of fundamental research and drug development, including the burgeoning field of RNA therapeutics.” Yin also is a leader of the Wyss Institute’s Molecular Robotics Initiative and Professor in the Department of Systems Biology at HMS.
ROCK is breaking the current limits of RNA structural investigations and enables 3D structures of RNA molecules to be unlocked that are difficult or impossible to access with existing methods, and at near-atomic resolution. We expect this advance to invigorate many areas of fundamental research and drug development, including the burgeoning field of RNA therapeutics.
PENG YIN
Gaining control over RNA
Yin’s team at the Wyss Institute has pioneered various approaches that enable DNA and RNA molecules to self-assemble into large structures based on different principles and requirements, including DNA bricks and DNA origami. They hypothesized that such strategies could also be used to assemble naturally occurring RNA molecules into highly ordered circular complexes in which their freedom to flex and move is highly restricted by specifically linking them together. Many RNAs fold in complex yet predictable ways, with small segments base-pairing with each other. The result often is a stabilized “core” and “stem-loops” bulging out into the periphery.

“In our approach we install ‘kissing loops’ that link different peripheral stem-loops belonging to two copies of an identical RNA in a way that allows a overall stabilized ring to be formed, containing multiple copies of the RNA of interest,” said Di Liu, Ph.D., one of two first-authors and a Postdoctoral Fellow in Yin’s group. “We speculated that these higher-order rings could be analyzed with high resolution by cryo-EM, which had been applied to RNA molecules with first success.”
Picturing stabilized RNA
In cryo-EM, many single particles are flash-frozen at cryogenic temperatures to prevent any further movements, and then visualized with an electron microscope and the help of computational algorithms that compare the various aspects of a particle’s 2D surface projections and reconstruct its 3D architecture. Peng and Liu teamed up with Liao and his former graduate student François Thélot, Ph.D., the other co-first author of the study. Liao with his group has made important contributions to the rapidly advancing cryo-EM field and the experimental and computational analysis of single particles formed by specific proteins.
“Cryo-EM has great advantages over traditional methods in seeing high-resolution details of biological molecules including proteins, DNAs and RNAs, but the small size and moving tendency of most RNAs prevent successful determination of RNA structures. Our novel method of assembling RNA multimers solves these two problems at the same time, by increasing the size of RNA and reducing its movement,” said Liao, who also is Associate Professor of Cell Biology at HMS. “Our approach has opened the door to rapid structure determination of many RNAs by cryo-EM.” The integration of RNA nanotechnology and cryo-EM approaches led the team to name their method “RNA oligomerization-enabled cryo-EM via installing kissing loops” (ROCK).
To provide proof-of-principle for ROCK, the team focused on a large intron RNA from Tetrahymena, a single-celled organism, and a small intron RNA from Azoarcus, a nitrogen-fixing bacterium, as well as the so-called FMN riboswitch. Intron RNAs are non-coding RNA sequences scattered throughout the sequences of freshly-transcribed RNAs and have to be “spliced” out in order for the mature RNA to be generated. The FMN riboswitch is found in bacterial RNAs involved in the biosynthesis of flavin metabolites derived from vitamin B2. Upon binding one of them, flavin mononucleotide (FMN), it switches its 3D conformation and suppresses the synthesis of its mother RNA.

“The assembly of the Tetrahymena group I intron into a ring-like structure made the samples more homogenous, and enabled the use of computational tools leveraging the symmetry of the assembled structure. While our dataset is relatively modest in size, ROCK’s innate advantages allowed us to resolve the structure at an unprecedented resolution,” said Thélot. “The RNA’s core is resolved at 2.85 Å [one Ångström is one ten-billions (US) of a meter and the preferred metric used by structural biologists], revealing detailed features of the nucleotide bases and sugar backbone. I don’t think we could have gotten there without ROCK – or at least not without considerably more resources.”
Cryo-EM also is able to capture molecules in different states if they, for example, change their 3D conformation as part of their function. Applying ROCK to the Azoarcus intron RNA and the FMN riboswitch, the team managed to identify the different conformations that the Azoarcus intron transitions through during its self-splicing process, and to reveal the relative conformational rigidity of the ligand-binding site of the FMN riboswitch.
“This study by Peng Yin and his collaborators elegantly shows how RNA nanotechnology can work as an accelerator to advance other disciplines. Being able to visualize and understand the structures of many naturally occurring RNA molecules could have tremendous impact on our understanding of many biological and pathological processes across different cell types, tissues, and organisms, and even enable new drug development approaches,” said Wyss Founding Director Donald Ingber, M.D., Ph.D., who is also the Judah Folkman Professor of Vascular Biology at Harvard Medical School and Boston Children’s Hospital, and the Hansjörg Wyss Professor of Bioinspired Engineering at the Harvard John A. Paulson School of Engineering and Applied Sciences.
The study was also authored by Joseph Piccirilli, Ph.D., an expert in RNA chemistry and biochemistry and Professor at The University of Chicago. It was supported by the National Science Foundation (NSF; grant# CMMI-1333215, CCMI-1344915, and CBET-1729397), Air Force Office of Scientific Research (AFOSR; grant MURI FATE, #FA9550-15-1-0514), National Institutes of Health (NIH; grant# 5DP1GM133052, R01GM122797, and R01GM102489), and the Wyss Institute’s Molecular Robotics Initiative.
reference link : https://wyss.harvard.edu/news/nanotechnology-enables-visualization-of-rna-structures-at-near-atomic-resolution/



{kind=link}